






目的:岛叶位于外侧裂底部,是肿瘤和血管畸形等病理过程的潜在发生部位。了解岛叶的解剖结构和血供对于在此区域进行精确的显微外科手术至关重要。
方法:本研究对20具人体尸脑(共40个半球)的岛叶动脉供血情况进行了研究。为增加可见度,向脑动脉灌注红色乳胶,并在手术显微镜辅助下进行解剖。
结果:供应岛叶的动脉平均数为96条(范围77–112条)。其平均直径为0.23毫米(范围0.1–0.8毫米),每条动脉的起源均可追溯至中大脑动脉(MCA),主要源自M2段。在22个半球(55%)中,有1至6条岛叶动脉起源于中大脑动脉的M1段,并供血于岛叶边缘区。在另外10个半球(25%)中,有1至2条岛叶动脉起源于中大脑动脉的M3段,并供血于上岛环沟或下岛叶环沟区域。岛叶动脉主要供血于岛叶皮层、极外囊,偶尔也供血于屏状核和外囊,但不供血于壳核、苍白球或内囊,这些区域由外侧豆纹动脉(LLAs)供血。然而,每个半球平均有9.9条(范围4–14条)岛叶动脉,主要位于后岛叶区域,其形态类似于穿通动脉,其中部分动脉供血于放射冠。还观察到较大、更突出的岛叶动脉(岛叶盖部动脉)(每个半球平均3.5条,范围1–7条)。这些动脉横越岛叶表面,随后向外侧环绕,并向盖部内侧表面延伸分支。
结论:全面掌握岛叶相关的复杂血管分布模式,以及精通岛叶解剖学,是制定恰当手术方案的先决条件,也是最终成功探查并切除该区域病变的必要基础。
岛叶(又称雷尔岛)构成外侧裂的基底,是覆盖脑胼胝体和基底核的大脑皮质内陷部分。要充分观察岛叶,必须沿其全长打开外侧裂。岛叶是边缘系统旁结构之一,被称为中间皮层,在解剖学和功能上位于旧皮层与新皮层之间。岛叶长期以来一直是研究和推测的对象;然而,其确切的功能范畴至今仍未完全明晰。它曾被描述为内脏感觉区、内脏运动区、补充运动区、前庭区,以及与言语和/或语言某些方面相关的区域。虽然已有概述岛叶某些特征的广泛概念,但明确界定其解剖连接的定义以及关于其功能的确切结论仍有待进一步研究。岛叶可能发生病理过程,例如神经上皮和脑膜肿瘤以及血管畸形;因此,了解岛叶的解剖结构和血管分布对于在此区域进行精细手术至关重要。
岛叶的解剖
作者在先前的文章中已详细描述了岛叶的解剖位置,因此本文仅作简要概述。岛叶呈三角锥形,由前、上、下岛周沟与盖叶分隔。环上岛沟位于额顶盖下,环下岛沟位于颞盖下,而前岛沟则位于眶额盖下。岛阈是进入岛叶的入口,并构成外侧裂谷(或前穿孔质)的外侧界限。岛中央沟自环上岛沟延伸至岛阈,将岛叶分为前部和后部两部分。前岛叶由前、中、后短岛回,以及其前下方的副岛回和横岛回构成。前岛叶的回融合形成岛叶顶端,这是其最表浅的区域。后岛叶由前、后岛长回组成,两者之间由后中央岛回沟分隔。岛叶构成覆盖在屏状核和豆状核上的皮层。
大脑中动脉的分段
大脑中动脉是所有脑血管中最复杂的(图1)。该动脉分为五个主要分段:M1至M5。M1(蝶骨嵴)段延伸自内颈动脉分叉处至大脑中动脉主干分叉处,后者位于岛阈部附近。M2(岛叶)段延伸自主分叉处至环岛沟,而M3(盖叶)段延伸自环岛沟至脑侧面(位于Sylvian裂内)。M4(脑回旁)段位于大脑的脑回旁表面,而M5(末端)段则构成M4段的远端延伸部分。
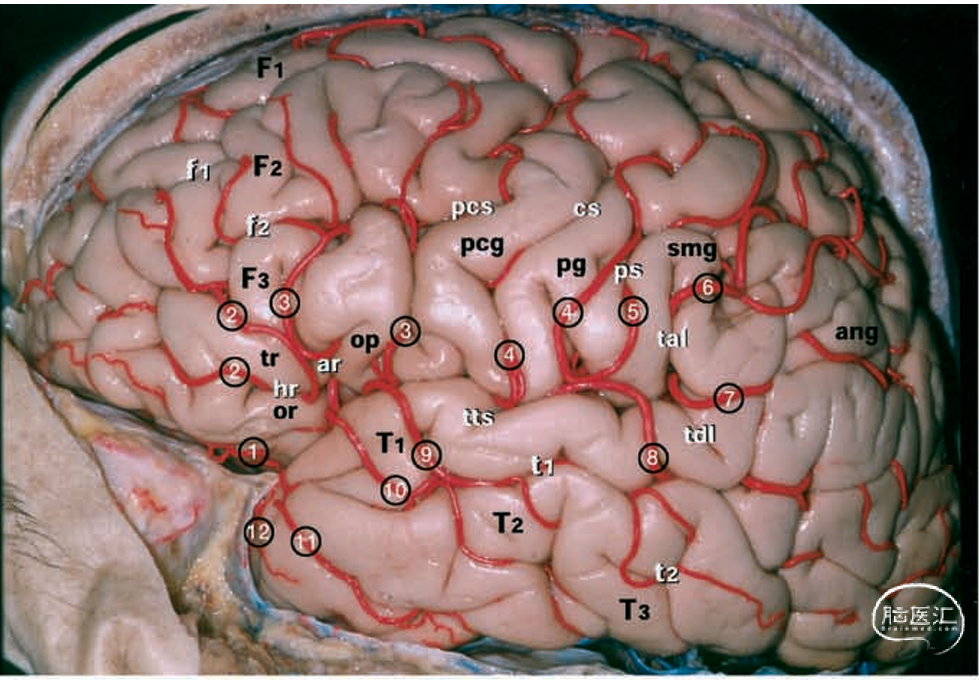
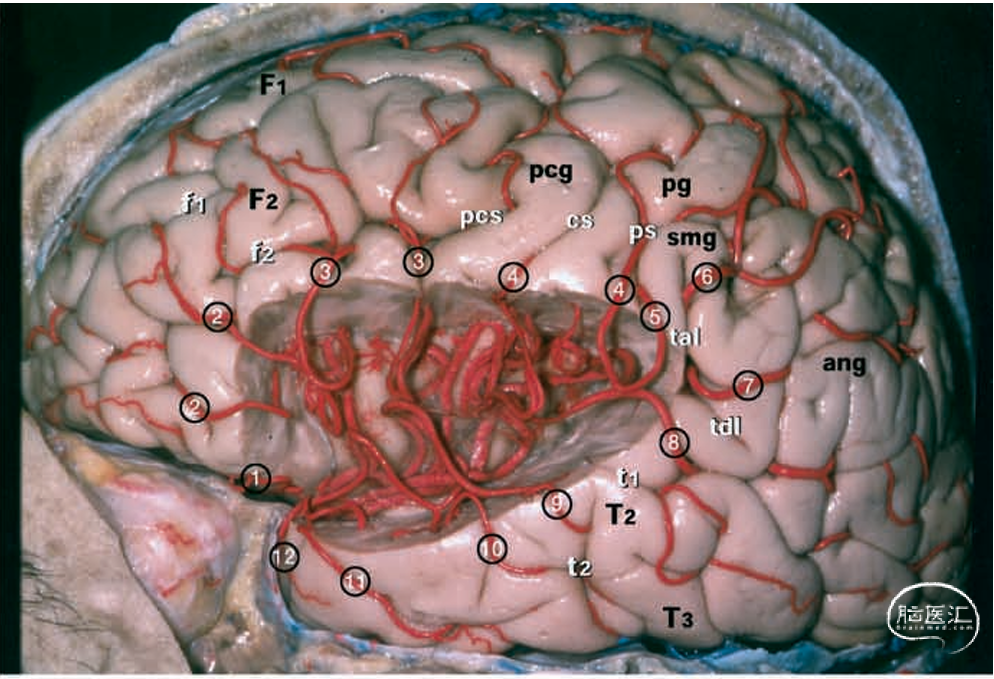
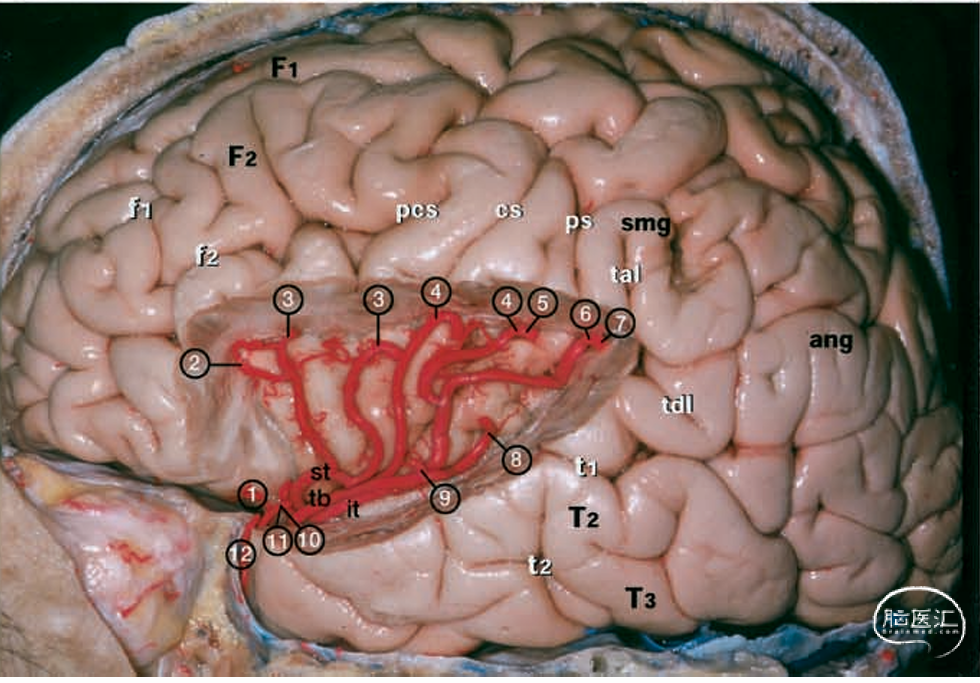
图1.大脑标本照片。上:左半球的外侧面由大脑中动脉(MCA)的M4和M5段供血。中:同一标本在切除全部盖部后,大脑中动脉的所有分支均得以保留。下:同一标本。中大脑动脉的M3、M4和M5分支已被切除,而供血于岛叶的M2分支则保留在岛叶皮层上方。白色字母表示脑回和脑裂。1=外侧眶额动脉;2=前额动脉;3=前中央动脉;4=中央动脉;5=前顶动脉;6=后顶动脉;7=角动脉;8=颞枕动脉;9=后颞动脉;10=中颞动脉;11=前颞动脉;12=颞极动脉;ang=角回;ar=侧裂上升支;cs=中央沟;F1=额上回;F2=额中回;F3,=额下回;f1=额上沟;f2=额下沟;hr=侧裂水平支;it=M2段的下干;op=F3的盖部;or=F3的眶部;pcg=中央前回;pcs=中央前沟;p=中央后回;ps=中央后沟;smg=缘上回;st=M2段的上干;T1=颞上回;T2=颞中回;T3=颞下回;t1=颞上沟;t2=颞下沟;tal=侧裂终末上升支;tb=大脑中动脉的颞支;tdl=侧裂终末下降支;tr=F3的三角部;tts=颞横沟。
材料与方法
本研究对20例福尔马林固定的尸头(共40个半球)进行了岛叶动脉的显微解剖研究。
先在颈部解剖并插管内颈动脉和椎动脉,随后用红色乳胶进行灌注以增强其可见度。颅骨切除后,小心切开硬脑膜,分离蛛网膜与软脑膜,并打开外侧裂。在手术显微镜下,采用6倍至40倍放大倍率,对LLAs的起始、管径、数量及走行进行了研究。由此可显露中大脑动脉(MCA)的皮质分支,并对其供血区进行分析。切除眶额叶、额顶叶及颞叶盖部后,显露了中大脑动脉的M1至M5段。在切除盖部过程中,特别注意保护覆盖岛叶表面的软脑膜。对岛叶动脉进行了研究,特别侧重于确定其起源位置,并测定其数量、直径及供血区域。在两具标本中,通过冠状切面观察来研究岛叶动脉的皮层下供血区。在三具标本中,我们采用纤维分离技术,界定了白质通路并分步进行分离,从而能够精确研究壳核、苍白球、前连合及内囊中的LLA供血区。
结 果
大脑中动脉的蝶骨段(M1)
在所有标本中,颈内动脉均在前穿孔质的中段分叉为中大脑动脉和大脑前动脉。大脑中动脉的M1段在侧裂深处、侧裂小谷内向外侧及上方走行,绕过岛叶边缘延伸至岛叶顶端,并在该处形成膝部。膝部的平均角度为97˚(范围90–130˚)。胫部距岛叶边缘的平均距离为4.8mm(范围2–9mm)。M1段的平均直径为3.21毫米(范围2.6–4毫米),平均长度为23.4毫米(范围15–38毫米)。M1段在侧裂谷内的走行方向可为前上外侧、上侧或后上外侧。区分M1段与M2段的界限是中大脑动脉的分叉,该分叉位于中大脑动脉膝部,紧邻岛叶边缘(图2)。我们将中大脑动脉的这一特定分叉命名为主分叉。在23个半球(57.5%)中,主分叉位于膝部。另有11个半球(27.5%)中,主分叉位于膝部远端4至10毫米处,而在剩余的6个半球(15%)中,主分叉位于膝部近端5至8毫米处。在40个半球中的2个(5%)中,M1段的颞支十分发达;在另一个半球(2.5%)中,额支十分发达,明显类似于中大脑动脉的主分叉;因此将其命名为“假分叉”。关于M1段及中大脑动脉主分叉部位的变异,已在第一作者(M.G.Y.)的先前文献中详细描述。观察到M1段分支遵循的解剖模式存在差异。根据其供血区域,可将其分为两组:皮质动脉和外侧豆纹动脉(LLAs)。
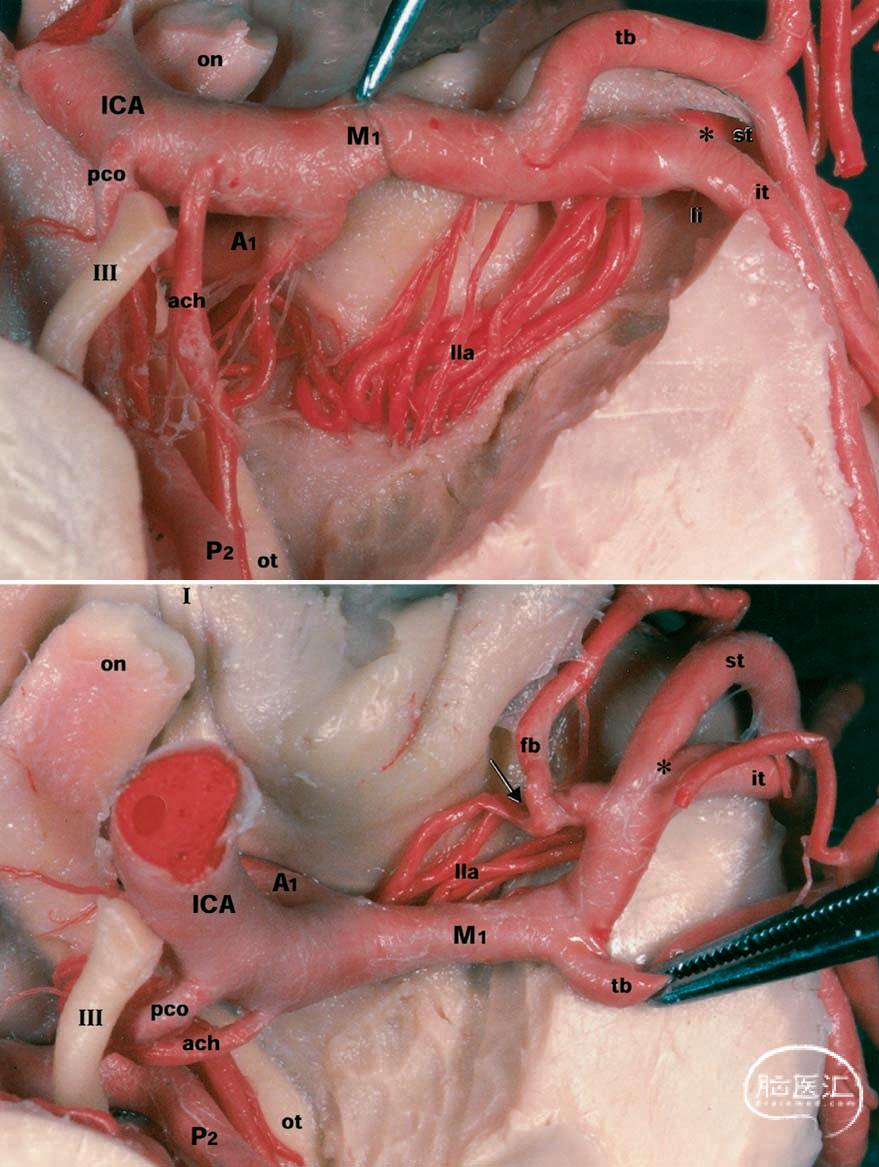
图2.大脑标本照片。上图:切除颞叶后,显示左侧中大脑动脉在前穿孔质区域的下视图。用镊子牵开M1段以显露外侧豆纹动脉(lla)。下图:抬起M1的颞支(tb)以旋转M1段,从而显露中大脑动脉的前支(fb)。箭头指示源自前支的外侧眶额动脉(lateral orbitofrontal artery)。星号指示大脑中动脉的主分叉。I=嗅神经;III=动眼神经;A1=前连通段;ach=前脉络膜动脉;li=岛叶阈;M1=大脑中动脉蝶骨段;on=视神经;ot=视束;P2=大脑后动脉脑池段;pco=后交通动脉。其他缩写请参见前一图的图例。
● 皮质动脉。在38个半球(95%)中,M1段分出1至3条主要皮质分支,这些分支主要位于M1段的外侧(75.8%),供血于颞叶;或位于该段的内侧(24.2%),供血于额叶。在7个半球(17.5%)中,我们发现了一条,偶尔两条直径为0.3至0.5毫米的小动脉(钩回动脉),供血于梨状皮质。我们未将此归类为主要皮质动脉。M1段主要皮质分支的变异被分为四种类型(图3)。在A型中,M1段分出一个颞(外侧)支。这一现象见于23个半球(57.5%);在17个半球(42.5%)中,存在一个颞支;在5个半球(12.5%)中,存在两个颞支;在1个半球(2.5%)中存在3条颞支。在B型中,M1段分出颞(外侧)支和额(内侧)支。这一现象见于14个半球(35%);在11个半球(27.5%)中,各有一条颞支和额支;而在3个半球(7.5%)中,有两条颞支和一条额支。在C型中,M1段仅分出一条额支,无颞支。这一现象见于1个半球(2.5%)。在D型中,M1段除外侧丘纹状动脉(LLAs)和钩回动脉外,不产生其他主要皮质分支。这一现象见于两个半球(5%)。
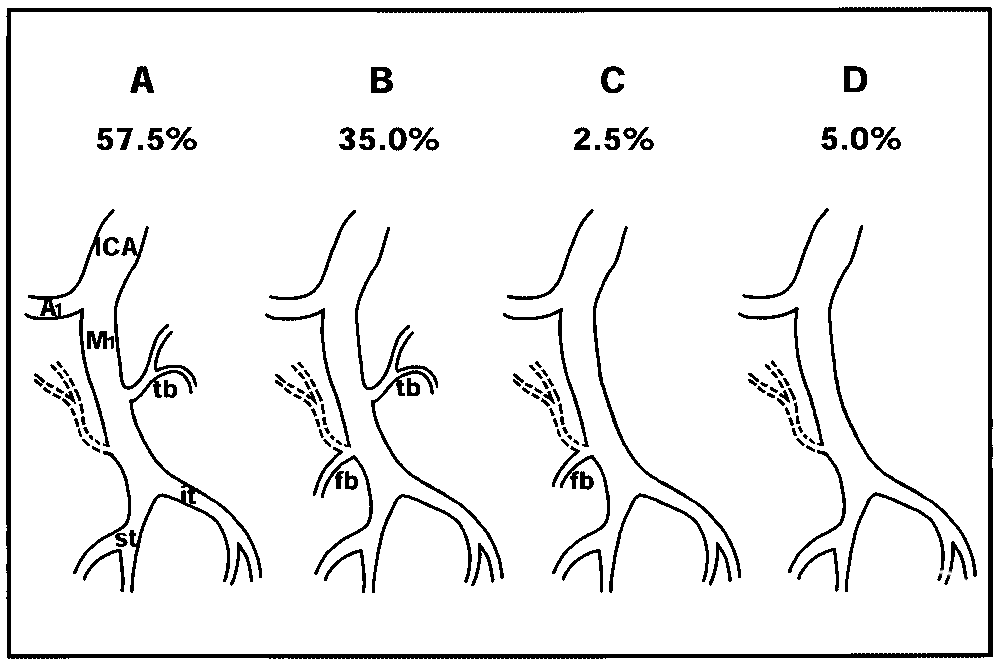
图3.显示M段皮质分支解剖变异的示意图。
A:M1段在57.5%的大脑半球中分出颞支。B:在35%的大脑半球中,M1段分出了颞动脉和额动脉分支。C:在2.5%的大脑半球中,M1段分出额支。D:在5%的大脑半球中,M1段未分出主要皮质分支(仅有LLAs和钩回动脉)。LLAs用虚线表示。A1前交通段(ACA的前交通段)。
● 外侧丘纹状动脉。在所有半球中,M1段均分出外侧丘纹状动脉,这些动脉主要位于M1段的下内侧面,位于侧裂小谷内。这些动脉穿行至前穿孔质的中央和外侧部分,在无名质的血供中起着重要作用,同时还供应壳核、苍白球、尾状核头和体、内囊及相邻的放射冠,以及前连合的外侧部分。每个半球内LLAs的数量在1至15条之间(平均7.75条),且M1段的长度与LLAs的数量之间似乎不存在明显的关联。LLAs在起源处的直径范围为0.1至1.5毫米(平均0.45毫米)。绝大多数(73%)LLAs的直径小于0.5毫米。其余(27%)直径较大,其中大部分分支成许多较小的分支。我们在每个半球中观察到至少一条、最多四条LLAs。LLAs的起源各不相同。78%起源于M1段,通常位于下内侧面。然而,在18%的病例中,LLAs起源于额叶或颞叶分支;在4%的病例中,LLAs起源于M2段的上干或下干,且位于中大脑动脉(MCA)主分叉附近。LLAs各分支之间在蛛网膜下腔内无交通。在15个半球(37.5%)中,M1段分出前支(外侧眶前动脉),而在9个半球(22.5%)中,该前支分出强有力的LLAs(图2下部)。在其余6个半球(15%)中,LLAs直接起源于M1段,靠近额支的起始处。从M1段分支后,LLAs突然转向,在起始处形成锐角,向内侧走行4至5毫米,随后向上转向进入前穿孔质的侧部。从侧面观察,LLAs呈现扇形分布,始于脑底,向外辐射,几乎覆盖整个内侧囊(图4)。
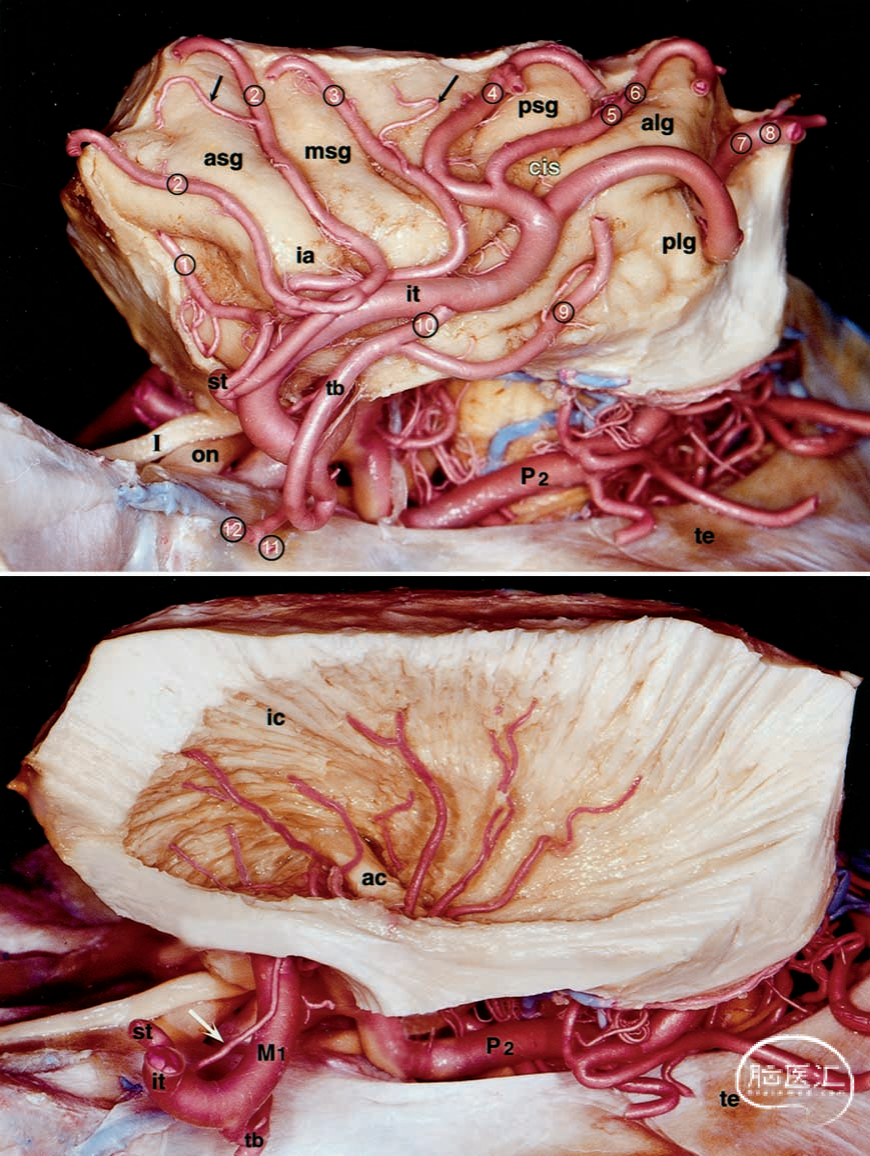
图4.大脑标本照片。上图:从岛周沟移除额叶、顶叶、枕叶和颞叶后,可见岛叶。岛叶的动脉起源于M2段。岛叶-盖叶动脉(前头所示)为岛叶和盖叶供血。下图:该区域的纤维分离显示,豆状核(已切除)的血供以及内囊由源自M1段的LLAS(箭头)供血。ac=前连合;alg=前岛长回;asg=前短岛回;cis=岛叶中央沟;ia=岛叶顶;ic=内囊;it=M2的下干;msg=中岛短回;P2=大脑后动脉的脑池段;plg=后岛长回;psg=后岛短回。其他缩写请参见前图图例。
在22个半球(55%)中,观察到M1段的小分支参与岛叶的供血。有1至6条动脉起源于M1段远端,并供血于岛叶边缘区域。在25个半球(62.5%)中,外侧眶额动脉起源于M1段,其分支延伸供血于横岛回和副岛回;该动脉演变为M3段,而M2段则缺如。同样地,当颞极动脉和前颞动脉起源于M1段外侧面的颞支时,这些动脉的M2段也缺如,且无分支供血至岛叶。
大脑中动脉(MCA)的岛叶(M2)段
观察到大脑中动脉通常在岛阈水平处分支为上干和下干,这两条干作为M2段走行于岛叶皮质上方。在35%的大脑半球中,上干比下干更粗;另有15%的大脑半球中两者粗细相等;而在剩余50%的大脑半球中,下干更粗。M2段上干的平均直径为2.51毫米(范围1.6–3毫米),M2段下干的平均直径为2.35毫米(范围1.3–3mm)。M2段上干与下干之间的夹角存在变异;平均值为91°(范围35–160°)。在3个半球(7.5%)中,M2段的上干或下干在分叉后立即分出一个或两个小的LLAs。
在22个半球(55%)中,M2段的上干或下干(以较占主导地位的干为准)在主分叉远端再次分叉,形成一个“中间干”。在18个半球(45%)中,该中间干源自上干;而在其余4个半球(10%)中,则源自下干。在5个半球(12.5%)中,这一二次分叉(中间干)发生在主分叉附近,给人以三叉分叉的印象。在另一个半球(2.5%)中,上干和下干均在主分叉点之后立即分叉,呈现四叉状。
在岛叶边缘区域,并非总是仅可见中大脑动脉的上干和下干;有时还会发现三至五条干动脉。沿上岛叶沟和下岛叶沟,M2段在过渡为M3段之前平均有9.6条分支(范围为8–12条分支)。这些分支主要起源于上干,并在前岛叶区域进一步分支。在每个半球中,前额叶动脉位于岛叶前点区域(图1、图4上部及图5)。前额叶动脉、前中央动脉和中央动脉,以及在22.5%的半球中,前顶叶动脉和后顶叶动脉,均从上干向岛叶呈扇形分布。它们主要供血于岛叶的前部。当到达上岛周沟时,这些分支急剧转折,形成M3段,即所谓的“烛台动脉”。
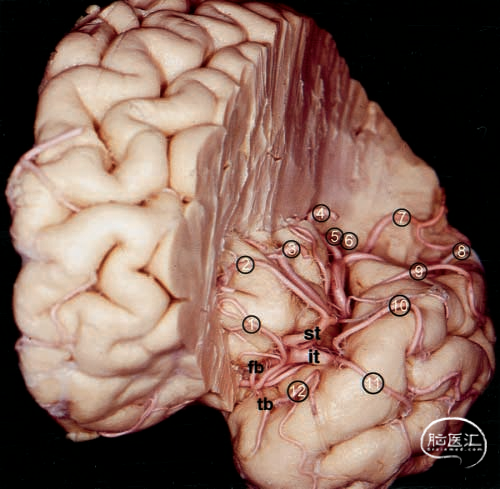
图5.切除额眶盖和额顶盖后左脑半球的前视图。位于岛环沟前上方的大脑中动脉上干M3段已被切断。供血于颞叶的下干得以保留。缩写说明参见前图图例。
在每个半球中,中央动脉(或包含中央动脉的干)部分或完全沿岛叶中央沟走行(图1、图4上部及图6)。从未观察到其起源于M1段分出的额叶或颞叶分支。岛叶中央沟是岛叶中血管分布最密集的区域。在后岛叶点区域,观察到一条或两条动脉行向后岛叶沟,形成M3段,随后在侧裂后方的脑皮质表面分叉,形成M4段。这些动脉分支代表了中大脑动脉沿侧裂的延续,随后延伸至岛叶后部,形成多条岛叶动脉。后顶动脉始终见于这些分支(图1和图4上)。中央动脉在三个半球中见于这些分支,而在另外四个半球中则见于前顶动脉、后顶动脉、角动脉和颞枕动脉。后颞动脉和中颞动脉起源于M2段的下干,并沿下岛周沟区域走行,最终形成M3段。颞支起源于M1,其走行方向始终位于其他M2段的侧方,而这些M2段位于岛叶表面。
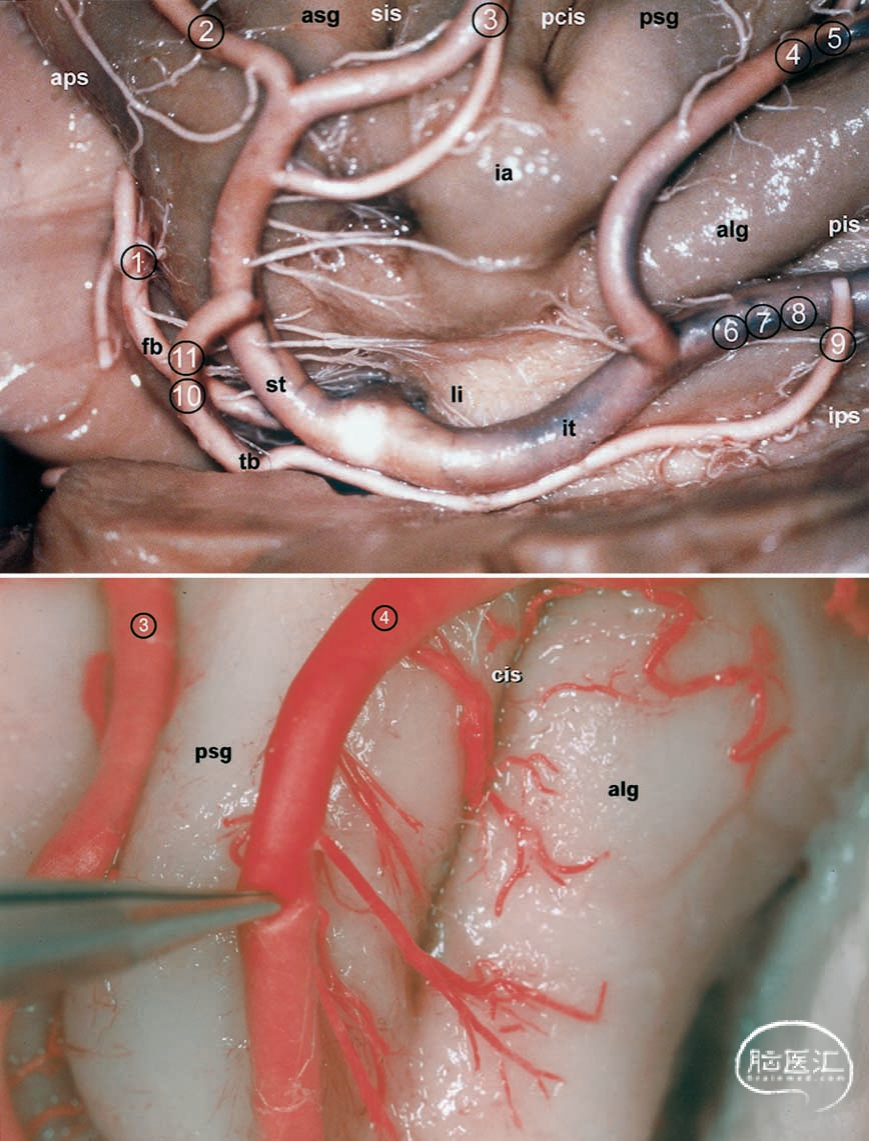
图6.大脑标本照片。上图:左侧岛叶的动脉。额支和颞支分别在前岛叶周围沟和下岛叶周围沟区域为岛叶供血。下图:中央动脉向中央岛回沟提供分支。aps=前岛回沟;ips=下岛回沟;pcis=前中央岛回沟;pis=后中央岛回沟;sis=短岛回沟。其他缩写请参见前图图例。
大脑中动脉的盖部(M3)段
M3段动脉束的走行始于岛叶周围的前、上、下沟,沿盖部的隐蔽内侧面延伸,并在外侧裂表面转为M4段的走行(图5)。M3段平行于M2段跨越岛叶,但延伸方向相反。M3段供血于盖叶的内侧面;然而,在10个半球(25%)中,它们还分出1至2条小岛内动脉,这些动脉供血于上岛环沟或下岛环沟区域。外侧眶额和颞极动脉表现出一个特殊特征:当这些动脉起源于M1段时,它们立即转变为M3段,不向岛叶发出分支。
大脑中动脉的侧裂旁段(M4)和终末段(M5)
M3段向外侧走行并穿出侧裂,在大脑半球外表面延续为M4段。M4和M5段由12条主要动脉组成,早期的出版物已根据其供血区域对这些动脉进行了记录和命名。这些动脉包括:外侧眶额动脉、额前动脉、中央前动脉、中央动脉、顶前动脉、顶后动脉、角回动脉、颞枕动脉、颞后动脉、颞中动脉、颞前动脉和颞极动脉(图1)。未观察到M4和M5段有任何分支供应岛叶。
岛叶的血液供应
岛叶的血液供应主要来自M2段。对40个半球的检查显示,有75至104条岛叶动脉起源于该段。然而,在22个半球(55%)中,有1至6条岛叶动脉起源于远端M1段,并供应岛阈(limen insulae)区域。在10个半球(25%)中,有1或2条岛叶动脉起源于M3段,并供应上岛周沟或下岛周沟区域。我们未观察到来自M4和M5段的任何分支供应岛叶。
在每个半球中,平均发现96条岛叶动脉(范围77–112条)供应岛叶(图4上部,图6和图7)。这些动脉的平均直径为0.23毫米(范围0.1–0.8毫米)。每个半球中平均有9.9条岛叶动脉(范围4–14条)类似于穿支动脉,其分布偶尔可延伸至放射冠(图7和图8)。我们有时观察到一条管径较大的岛叶动脉,它沿岛叶表面走行,然后向外侧弯曲,发出分支同时供应岛叶和岛盖的内侧面。我们将这条动脉命名为“岛盖动脉”(insuloopercular artery)(图4上部)。
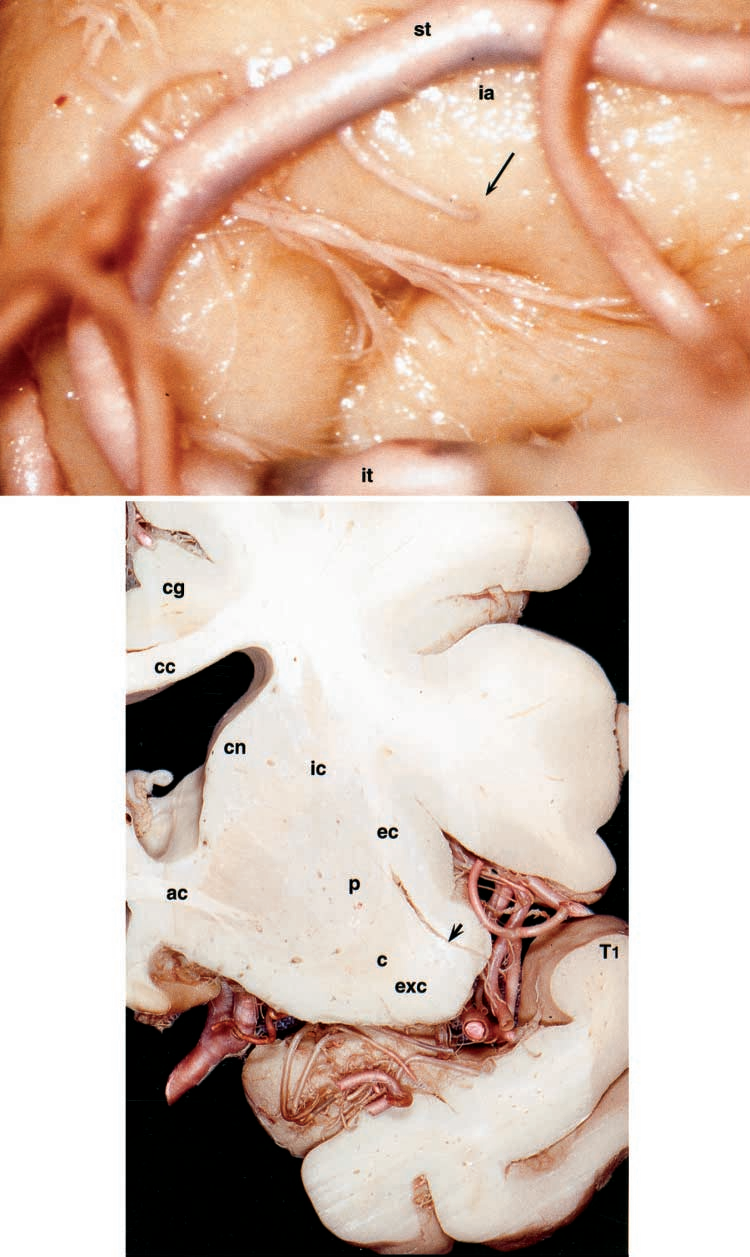
图7.大脑标本照片。上图:穿通样岛叶动脉(箭头)供血于岛叶顶端(ia)。下图:显示左侧半球经侧裂谷的冠状切面穿通样岛叶动脉(箭头)起源于M2段,供血于岛叶皮层、极外囊(exc)、胼胝体(cc)及外囊(ec)。cc=胼胝体;cg=扣带回;cn=尾状核;P=壳核。其他缩写请参见前图图例。
大约85%至90%的岛叶动脉较短,仅供应岛叶皮质和极外囊;10%为中等大小,还供应屏状核和外囊;其余3%至5%较长,可延伸至放射冠(图8)。长岛叶动脉呈穿支样,主要位于岛叶的后部区域。壳核、苍白球和内囊由豆纹动脉(LLAs)供血(图4下部)。研究发现,外囊是豆纹动脉和岛叶动脉供血区域的分界。我们未观察到岛叶动脉与豆纹动脉之间存在明显的大体交通支。
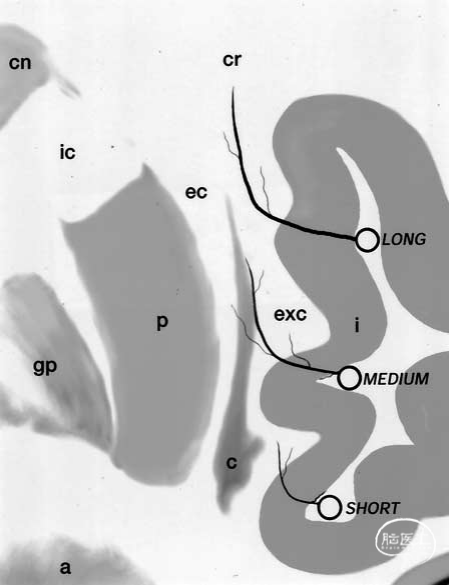
图8.示意图显示,约85%至90%的岛叶动脉较短,主要供血于岛叶皮层(i)和外囊;10%为中等长度,除上述区域外,还供血于胼胝体和外囊;剩余的3%至5%较长,其走行可达放射冠(cr)。a=杏仁核;gp=苍白球;P=壳核。其他缩写请参见前图图例。
讨 论
大脑中动脉(MCA)是脑血管中最复杂的血管。该动脉供应几乎整个大脑半球的外侧面,以及岛叶、豆状核和内囊。许多研究已对MCA的显微外科解剖,尤其是M1段和豆纹动脉(LLAs)的解剖,进行了详细的检查和分析。众所周知,慢性高血压会引起脑血管的病理改变,导致血管闭塞或破裂,进而分别引发腔隙性脑梗死或脑内出血。自发性脑内出血最常见的部位是豆状核区,见于35%至50%的患者。由于豆状核结构位于豆纹动脉和赫布纳动脉(Heubner's artery)的供血区域内,当怀疑出血源于这些血管时,熟悉豆纹动脉的解剖结构显然至关重要。
Marinković及其同事观察到豆纹动脉的数量在3至18条之间(平均9条)。他们探查并描述了每一条豆纹动脉的起源,发现这些血管在其脑外段或脑内段均无相互交通。Duret曾提出,豆纹动脉向外侧走行可达壳核。此外,他断言更外侧的结构,如屏状核和外囊,其血液供应来自穿入岛叶的血管。Beevor赞同Duret的观点,确认了豆纹动脉的这一外侧边界。两者均指出,外侧豆纹动脉与岛叶动脉之间不存在交通。Shellshear在结扎M2段后,向大脑中动脉注入染色剂。他发现染色剂出现在纹状体、屏状核和外囊中,而岛叶皮质则清晰无染色。我们的研究证实,外囊是豆纹动脉和岛叶动脉供血区域的分界线。我们未发现这两套动脉系统之间存在明显的大体交通证据。岛叶动脉可以被电凝,以使岛叶区的内在肿瘤和血管畸形去血管化,同时不会损害壳核和内囊的血供。然而,由于可能导致放射冠梗死(图8),应保留岛叶长动脉。有观察发现,某些纹状体动静脉畸形(AVM)由这两组动脉共同供血,这表明豆纹动脉与岛叶动脉之间可能存在微小的交通支(图9)。
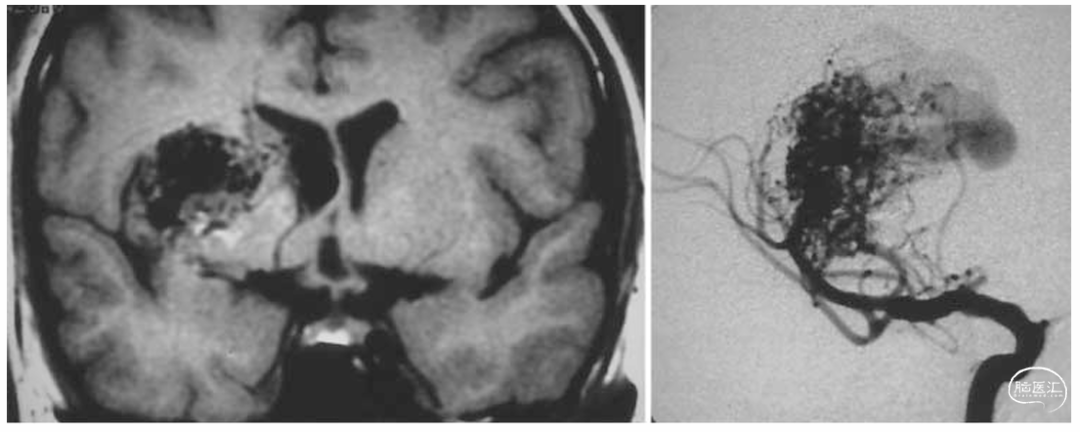
图9.左图:冠状位T1加权MRI显示纹状体区域的异质性病变(动静脉畸形,AVM)。右图:右侧ICA血管造影的前后投影视图显示纹状体AVM。注意AVM的供血血管起源于岛叶动脉,这些动脉来自M2段和LLA。
额支起源于M1段,具有重要的外科意义。在两个半球(5%)中,豆纹动脉(LLAs)起源于M1段的额支(图2下部)。位于M1段该区域的动脉瘤可能被误诊为大脑中动脉(MCA)分叉部动脉瘤。手术中,若在此处对动脉瘤施以夹闭,可能会闭塞那些起源于额支的豆纹动脉,这些豆纹动脉的起源往往靠近M1段上额支的起始处。这一操作可能导致患者术后出现偏瘫。必须认识到,豆纹动脉也可能起源于靠近MCA分叉处,偶尔也可起源于M2段的上干或下干。术前和术中充分考虑这些血管与动脉瘤的确切位置及关系至关重要。通过详细的血管造影可以确定豆纹动脉及相关动脉的形态,并在手术探查中验证其位置。
Umansky等人描述了MCA近端节段的详细解剖结构。他们的研究证实,微血管重建手术(如吻合术、移植术和动脉分支再植术)在岛叶区域是可行的。他们观察到,将M2段抬起3至5毫米并不会牵拉供应岛叶的软脑膜血管。他们还指出,M2段具有进行微血管吻合的潜力。然而,充分了解M4段的哪条皮层分支起源于M2段的哪个主干至关重要。特别需要调查的是哪个M2主干供应中央区。我们的观察显示,中央动脉起源处的主干总是沿岛中央沟的部分或全部走行。顶前动脉、顶后动脉和角回动脉的起源和走行也表现出特殊特征:两者均跨越长岛回到达岛后点,并作为M3段的一部分,继续在侧裂深处的岛后沟内走行。血管造影显示这些动脉具有独特的形态,呈现出MCA向后延伸的外观。
我们在确定M1段与M2段的连接处以及MCA主分叉的位置时遇到了困难。根据文献记载,这一问题仍存在争议,导致关于M1段的真实模式和测量产生混淆。在我们的样本中,7.5%的半球显示出M1段的额支或颞支非常粗大,它们类似于MCA的分叉,从而阻碍了对其真实位置的明确识别。Yasargil将这种变异称为“假性分叉”。由于豆纹动脉典型地起源于M1段,因此定位豆纹动脉及其起源点对于区分MCA的主分叉具有重要意义。
用于描述MCA二分叉、三分叉或四分叉的术语引发了进一步的混淆。我们认为MCA存在一个主分叉,但希望补充我们的发现并对两种观察结果进行评论。在12.5%的半球中,中间干经过主分叉附近,给人造成MCA三分叉的印象。在2.5%的半球中,上干和下干在主分叉后立即再次分叉,给人造成MCA四分叉的印象。在Umansky等人检查的尸体标本中,作者观察到66%的半球为二分叉,26%为三分叉,4%为四分叉。Gibo等人在其尸体研究中观察到,78%的半球为二分叉,12%为三分叉,10%分为多个主干。
在我们的标本中,未观察到副大脑中动脉、MCA重复或M1段窗孔畸形。几位作者曾报道观察到副大脑中动脉,它起源于大脑前动脉(ACA),走行路径与赫布纳动脉(Heubner's artery)相似,很少发出穿支。然而,已观察到有皮层分支供应额叶眶面外侧部分。根据多位作者的报告,这种变异见于0.3%至3%的半球。“MCA重复”(即两条MCA成对起源于颈内动脉)这一术语可能由Teal等人首次提出,随后出现在其他文献中。文献中也有关于M1段窗孔畸形的描述。
结 论
在本研究中,我们详细探查并研究了岛叶复杂的血管分布。我们致力于定义、描述并阐明该区域错综复杂的血管模式和各种动脉通路,并结合该区域的显微外科解剖学知识进行阐述。此举旨在将这些知识整合、协调并应用于手术规划及病理性病变切除的手术过程中。
Ali Krisht教授是位备受尊敬的神经外科学教授,服务于美国小石城CHI St.Vincent North医院。作为阿肯色神经外科研究院(ANI)的院长和首席神经外科医生,他在脑血管病、垂体瘤和颅底肿瘤领域发表了系列经典论著。


Ali Krisht教授在海绵窦、脑动脉瘤和胶质瘤手术治疗方面作出的开创性贡献,为他赢得了多个奖项,其中包括2019年的Herbert Olivecrona奖(被誉为神经外科的诺贝尔奖)。Castle Connolly和“当代神经外科”均将他评为美国顶尖1%的神经外科专家。毫无疑问,他是先进显微神经外科领域的杰出领袖之一。

脑医汇-神外资讯有幸获得了对Ali Krisht教授经典著作进行中文翻译和传播的独家授权,将陆续通过连载发布,方便大家分享学习。
特别感谢ANI显微神经外科解剖室联合主任蔡理医生对Krisht教授经典著作的整理,同时感谢郑州大学第一附属医院神经外科李天豪副主任医师和复旦大学附属华山医院宋剑平主任医师在内容整理及组织编译、审校工作上的辛勤付出。
以上是第三十三期内容:《岛叶的动脉》。感谢郑州大学第一附属医院神经外科李天豪副主任医师,以及蔡理医生的编译校对。
译者

● 医学博士,郑州大学第一附属医院神经外科副主任医师,美国阿肯色州神经科学研究所(ANI)访问学者。专业方向:颅底肿瘤。

![]()
声明:脑医汇旗下神外资讯、神介资讯、神内资讯、脑医咨询、Ai Brain 所发表内容之知识产权为脑医汇及主办方、原作者等相关权利人所有。
投稿邮箱:NAOYIHUI@163.com
未经许可,禁止进行转载、摘编、复制、裁切、录制等。经许可授权使用,亦须注明来源。欢迎转发、分享。
