




纷繁复杂的动作执行和动作学习过程都需要中枢神经系统中各种神经网络的参与。其中,脑干和基底神经节是处理运动信息的关键,其中也都聚集着功能特异的神经元,基于不同轴突投射、突触输入,以及基因表达,这些神经元又分成不同的层次,由此实现环路解剖学与功能的对应关系。脑干和基底神经节中的神经元群形成了多步处理环节,专门用于执行特定的运动;然而,揭示这两个结构之间的连接和信息传递的相关研究才刚刚起步。
脑干和基底神经节也参与了更广泛的系统层次的网络。其中,重要的网络组成部分包括皮层投射神经元、小脑输出神经元和中脑多巴胺能神经元。特定的动作环路能够被增强、抑制,彼此之间协同工作或竞争,或者通过可塑性产生适应性行为。作者认为,运动系统中这种高度特定的环路组织是支持行为特异性的核心成分,同时也为行为的灵活性提供了充分的基础。
本篇重量级综述在这篇综述中,介绍了四部分内容;
在上节中,我们已详细阐述了第一部分:脑干的神经元如何参与两种与肢体有关的行为:
运动速度和方向性的调节;
在身体没有移动的情况下调控与前肢有关的各种运动。
鉴于综述篇幅,
AiBrain将本文分为四部分,
以专题的形式为读者带来详细的解读。
在这篇文章中,我们将重点介绍综述的第二部分:基底神经节环路的组织原则,重点阐述了与运动息息相关的直接通路/间接通路。
Part 2
基底神经节环路组织
上一节回顾的工作表明脑干神经元的特定细胞群参与不同的运动。这些细胞群接受来自上游结构的兴奋性输入(包括皮质和上丘)可能是它们激活的原因。然而,脑干神经元也接收来自基底节的直接和/或间接输出,最显著的是来自黑质网状部(SNr)的GABA能神经元,其本身是中脑的一部分(图1a)。脑干细胞群的基底节调节被认为对不同运动的选择和/或执行至关重要。
一个经典模型表明,对于特定运动的选择和执行,脑干细胞群需要兴奋性输入激活和基底节输出结构解除抑制。与该模型一致的是,在训练有素且有回报的环境中进行扫视眼球运动时存在抑制苍白球和上丘的SNr输入(图1b)。但是SNr神经元提供强直抑制的模型在运动过程中暂停,从而解除对下游脑干细胞群的抑制,这一点尚未得到广泛证实。
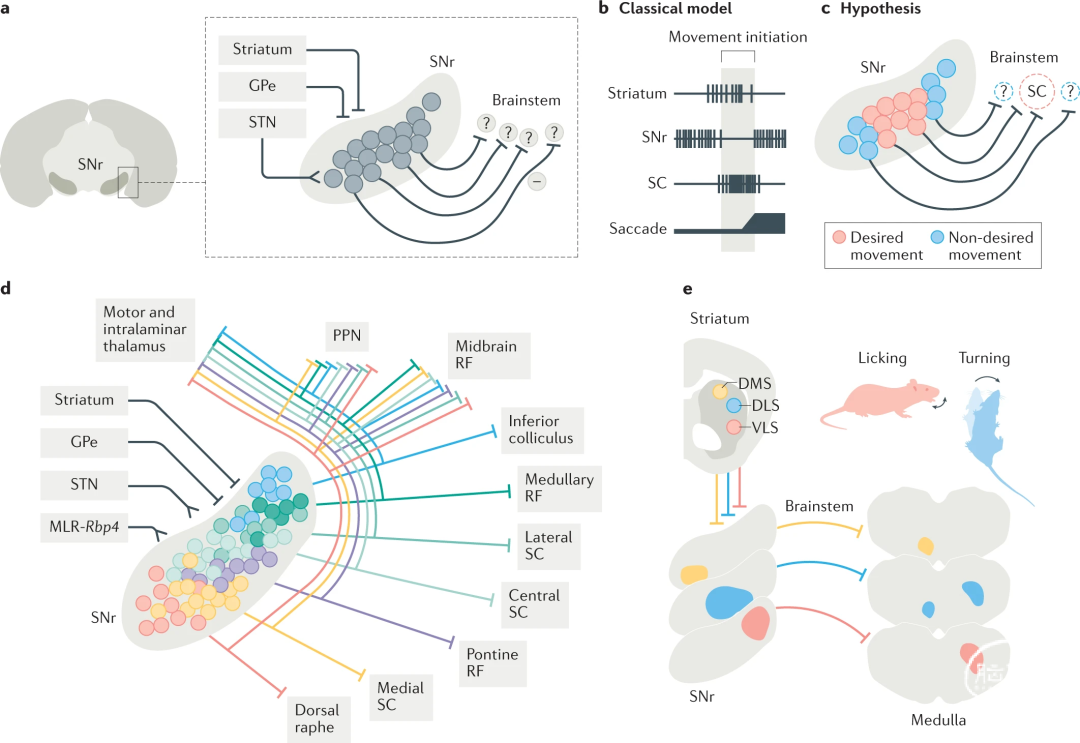
图1:基底节和脑干环路之间的相互作用。
a |黑质网状部(SNr)神经元是基底节和脑干运动中心之间的守门员。SNr神经元整合来自纹状体、丘脑底核(STN)和基底节内苍白球(GPe)的信息。它们的抑制性输出(-)如何与脑干相互作用尚不清楚(用?表示)。
b |经典模型提出,SNr神经元因上游纹状体神经元激活而暂停紧张性放电,导致上丘(SC)下游神经元去抑制。因此,兴奋性SC神经元上调放电并参与扫视的执行。
c |选择和抑制模型表明,相对于下游靶点,SNr神经元具有高度的组织性。根据该模型,只有与期望行为(红色)相关并选择性投射到特定下游靶点(SC)的SNr神经元会被抑制,但周围的SNr神经元(蓝色)不会被抑制并将继续活跃,从而抑制非期望行为。然而,到目前为止还没有经过实验测试。
d |不同的SNr神经元群投射到特定的脑干靶点(用不同的颜色显示),但除此之外,所有这些神经元都会产生轴突侧支到脚桥核(PPN)、中脑网状结构(RF)以及运动丘脑和层内丘脑。SNr神经元的兴奋性输入由三角形终端(STN和MLR-Rbp4神经元,由转基因Rbp4标记)显示,而抑制性输入由钝箭头(纹状体和GPe)显示。
e |来自不同纹状体区域的局部定位投射(背内侧纹状体(DMS),黄色;背外侧纹状体(DLS),蓝色;腹外侧纹状体(VLS),红色)进入SNr的不同区域。这些不同的SNr亚区也针对尾侧脑干的不同下游靶点,如纹状体到SNr的顺行跨突触病毒转移实验所示。刺激DLS和VLS中的纹状体投射神经元分别引起舔舐或转身行为,这与延髓中相应SNr神经元下游的功能专一性一致。
令人惊讶的是,虽然许多SNr神经元在运动过程中确实受到抑制,但其它神经元则被激活。这可以解释为,原则上SNr神经元可以同时针对谷氨酸能和GABA能脑干神经元,因此兴奋的神经元可能仍然会解除对下游运动中心的抑制。另一种可能性是,这种现象反映了一种中心环绕环路机制(图1c)。在这个中心-环绕模型中,被抑制的SNr神经元专门针对执行选定的期望运动所需的脑干细胞群,而被激活的周围SNr神经元投射到其它区域以抑制竞争性的非期望运动(图1c)。事实上,在细胞群水平上,运动过程中受到抑制的SNr神经元可以预测运动起始,而兴奋神经元则不能。要使该模型普遍适用于SNr输出信令,必须满足几个条件:
为了传达动作特异性,不同的SNr细胞群应该投射到与这些动作调控相关的不同脑干区域,而不是一般地传播到所有下游区域。
输入到SNr神经元的信号应该表现出特异性。例如,在特定运动中被抑制的SNr神经元应该从纹状体获得与该特定运动执行相关的特定输入,但周围的SNr神经元应该获得不同的输入,而不是被抑制。
接下来,作者将介绍这两个主题的实验进展。
最近的研究表明,不同的SNr细胞群确实针对脑干内的不同区域(图1d)。从脑干七个主要区域(延髓和脑桥网状结构、上丘、下丘和中缝背侧的三个中外侧不同区域)中的每一个逆行标记的SNr神经元彼此之间显示出稀疏的侧支循环(图1d),表明平行的SNr通道针对不同的脑干区域。在SNr范围内,这些不同的细胞群在显著的空间组织模式中被发现。而更多的前外侧SNr区域投射到上丘的不同区域,更多的后内侧区域投射到脑桥和髓质的尾部脑干区域(图1d)。
SNr亚群也表现出不同的电生理特性,这取决于投射靶点。多数的外侧SNr神经元体积大、放电快、膜时间常数快,而大多内侧SNr神经元放电速度慢、衰减常数低。
外侧和内侧SNr神经元也被认为表现出不同的功能,多数外侧神经元参与运动,多数内侧神经元参与低运动活动和低觉醒状态,如睡眠。
尽管不同的SNr细胞群优先针对某些脑干区域,但它们都向丘脑和中脑区域发送侧支,包括PPN和中脑网状结构,部分包含在MLR内(图1d)。然而,不同的SNr通道在丘脑内表现出空间分离,表明它们整合到空间上紧密但不同的神经元环路中:上丘投射的SNr神经元终止于丘脑的更多外侧区,而脑桥投射和髓质投射的SNr神经元则定位于丘脑的内侧束旁区和腹内侧区。这些发现表明,针对不同脑干区域的SNr亚群与特定的丘脑区域有联系。这可能导致参与相同运动的脑干和丘脑皮质环路同时受到调节,从而使这些输出通道特定于脑干靶点和丘脑的特定区域。
以丘脑为靶点的侧枝还可以将待执行运动的有效复制信号传递到丘脑和皮层,用于监测正在进行的运动或学习。SNr细胞群也可能以MLR中功能不同的神经元为靶点。之前的研究表明,MLR的SNr输入会影响姿势调整和运动参数。将这些发现与MLR亚群的最新发现结合起来,可以想象,这可能是通过与姿势相关的脊髓投射MLR神经元和功能上与运动相关的髓质投射神经元的特定相互作用实现的。一般来说, SNr输出通道组织的特异性表明,SNr可能与不同的靶点在功能上相互作用,以调节运动中的位移,这可以在未来进行实验测试。例如,不同的SNr细胞群可能会调节涉及运动(速度和方向)的特定脑干模块,而不是本文前面描述的熟练前肢运动。
理解SNr的突触输入组织对于确定不同SNr亚群在行为过程中的招募方式也至关重要(图1d),因为SNr神经元兴奋性和抑制性输入的精细平衡决定了它们的功能特性。SNr神经元接收来自纹状体和外侧苍白球(GPe)的抑制性输入,而兴奋性输入则由MLR的丘脑底核(STN)和Rbp4转基因阳性神经元提供(图1d)。Rbp4转基因阳性MLR神经元位于邻近PPN的mRT内侧,不同于下行兴奋性MLR细胞群(图1b)。它们在前肢行为(如处理和梳理)期间优先活跃,但在后腿直立和移动期间不活跃。光遗传干扰实验表明,这些神经元在各种身体运动中起着重要的调节作用。目前尚不清楚Rbp4转基因阳性MLR神经元是否针对特定的SNr神经元亚群,但此类信息已经可用于SNr的其它突触输入。
值得注意的是,基底神经节的纹状体输入在空间上是分离的,并携带不同的功能信息(图1e)。不同的纹状体区域针对不同的SNr区域,纹状体的背外侧区域针对更外侧的SNr,腹内侧纹状体区域终止于更内侧的SNr。最近的两项研究发现,纹状体以一种高度局部投射的方式通过高度组织的基质连接到基底节结构。具体而言,背内侧纹状体(DMS)、背外侧纹状体(DLS)和腹外侧纹状体(VLS)针对不同的SNr和GPe亚区,通过这种特定的投射模式,它们保持脑干和丘脑区域的良好联系(图1e)。第二项研究揭示了一个更精细的连接图谱,也显示了纹状体与SNr和GPe之间的组织差异。虽然GPe中似乎存在高度空间分隔的分布,但还是能观察到一些传入通路投射到SNr;目前尚不清楚这项研究的功能意义。也许与此相关的是,即使在背内侧纹状体和DLS向投射到上丘同一区域的SNr神经元提供输入的情况下,这些细胞群也针对不同的层,这表明靶点向下游网络的功能差异。
为了探索特定通路所携带的差异功能模型,光遗传刺激实验提供了第一个见解:刺激腹外侧纹状体(VLS)中的SNr投射神经元(该区域之前与口面部运动有关)可诱发舔舐行为,而刺激背外侧纹状体(DLS)中的类似神经元(之前与身体运动有关)可调节身体转动(图1e)。这些观察结果与以下事实一致:DLS受体SNr神经元包括指向脑干Gi区的投射,与方向调控有关,纹状体腹外侧受体SNr神经元也以与口面部运动有关的延髓中心为靶点。纹状体、SNr和下游运动中心之间的其它身体运动,包括运动和熟练的前肢运动,如何实现类似的功能联系仍有待确定(见下文),但已知的解剖学路线图提供了有价值的切入点。有趣的是,分离的基底节环路也与丘脑和皮质形成分离的闭合环路,这表明基底节输出通路可以调节自身的输入,并表明特定的运动相关信息在更广泛的网络中传递,而不仅仅是传输到脑干中心执行。
对纹状体投射整体解剖结构的深入了解作者提出了一个问题,即在一个纹状体区域内是否存在其他多样性?这可能有助于理解未来投射环路的功能。有趣的是,即使在一个局限的纹状体区域内,就躯体和运动表征而言,也可能存在高度的粒度,这可能不会反映在光遗传刺激实验中。在小鼠进行不同的身体运动(包括运动、方向变化和后腿直立)时,对DLS中的数百个纹状体投射神经元(SPN)进行成像的最新研究表明,不同的神经元群在不同的运动模式下是活跃的,单个SPN表现出高度的运动特异性(图2a)。此外,DLS中的神经元编码详细的任务相关运动。GABA能SPN也根据其靶点分为两大类(图2b,c)。纹状体可以通过直接纹状体投射(dSPN)和间接纹状体投射(iSPN)调节SNr的活性。dSPN与SNr神经元有直接的突触联系,并表达D1型多巴胺受体(图2b),调节神经元的兴奋性。相比之下,iSPN只能通过GPe和STN间接影响SNr神经元,并表达D2型多巴胺受体(图2c),后者对神经元兴奋性进行负性调节。这些发现证实了经典假设,即dSPN在运动过程中是活跃的,直接抑制基底节输出神经元,从而解除对下游运动中心的抑制,而iSPN在运动过程中是不活跃的。
根据本文前面提出的SNr的中心-环绕模型,作者认为dSPN解除运动对应的下游环路的抑制,而iSPN将积极抑制不必要的运动。有趣的是,DLS中的dSPN和iSPN在运动过程中都是活动的,它们的活动是特定于运动的。对单个神经元的成像显示,大多数dSPN或iSPN神经元在特定的行为群中是活跃的,并显示出类似的运动特异性(图2b,c)。
两个细胞群同时钙信号记录还显示了特定行为模式执行期间特定的dSPN与iSPN活动模式。来自dSPN和iSPNs的活体膜片钳膜电位记录显示,这些神经元类型在毫秒时间尺度上具有不同的去极化时间。此外,尽管dSPN活动直接到达SNr(图2b),但在到达SNr之前,其它信息可以在GPe和STN中间基底节处理站的间接通路中整合(图2c)。这些额外的处理步骤可以增加间接途径信息流的异质性。此外,最近的研究表明,虽然纹状体区域和GPe之间存在精细的局部分布,但纹状体似乎更加明显地对SNr直接输入。因此,为了更好地理解来自纹状体地动作特异性信息是如何在基底节环路中传递的,对间接通路不同区域的神经元活动进行记录是至关重要的。有趣的是,GPe和STN中的不同结构域和/或细胞类型表现出不同的运动功能,甚至在人类中也是如此。此外,GPe(而非SNr)将投射发送回纹状体(图2c),允许进一步处理。GPe本身就是一个输出区域,直接投射到皮质和丘脑(图2c)。因此,iSPN有机会通过抑制GPe向基底节以外的下游环路的投射来解除基底节外环路的抑制,就像dSPN一样。此外,应该考虑的是,即使在经典情况下,当iSPN的活动导致SNr神经元兴奋时,SNr抑制性输出神经元也可以针对运动的抑制性神经元,从而导致去抑制。因此,这些最近的研究需要仔细描述所谓的间接通路,以及投射到不同运动中心的基底节投射群的活动。
一些研究试图通过分析基因表达谱来确定dSPN和iSPN的功能特异性是否也反映在基因表达水平上。有趣的是,SPN的核心分子特征似乎在整个纹状体中都是保守的,但除此之外,一些基因的表达在纹状体区域或纹状小体或基质小室中存在差异。
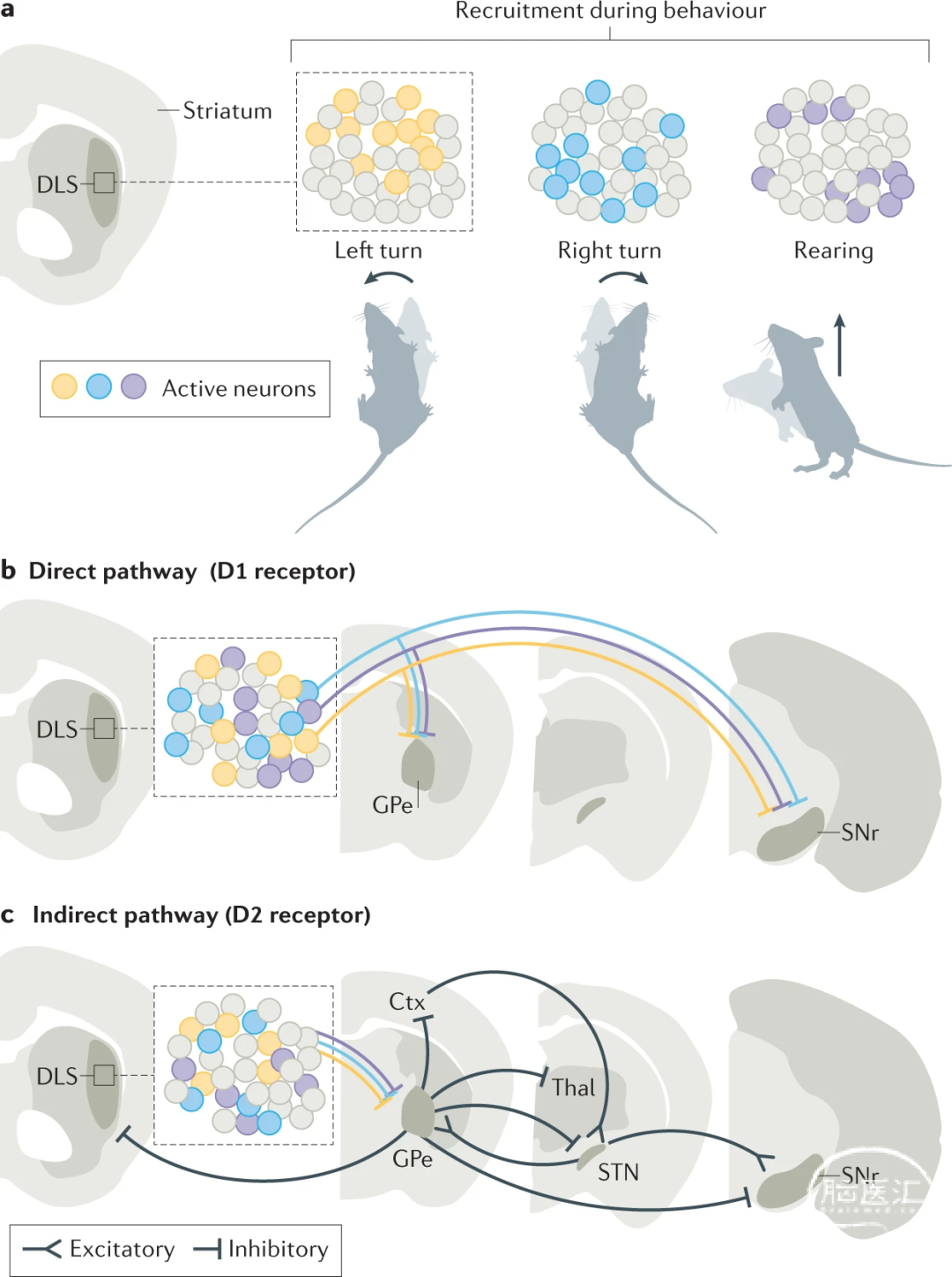
图2:纹状体环路的动作特异性。
a |使用背外侧纹状体(DLS;左)的钙成像监测纹状体投射神经元(SPN)活动表明,在不同的行为过程中,如左转、右转和后腿直立时,会招募不同的、特定于动作的神经元群。与这些行为相关的被招募神经元用彩色阴影表示,灰色表示未被所示行为招募的神经元。
b |包含直接通路(表达D1受体)的 SPN直接投射到黑质网状部(SNr),少量侧支则投射到外苍白球(GPe)。
c | 包含间接通路(表达D2受体)的SPN终止于GPe。许多区域是GPe神经元的靶点,部分是不同的神经元细胞群。其中包括纹状体本身、丘脑底核(STN)、SNr、皮质(Ctx)和丘脑(丘脑)。此外,STN也是间接通路的一部分,以GPe和SNr为靶点,自身接收皮质输入(超直接通路)。兴奋性投射显示为三角形末端,抑制性投射显示为钝箭头。
上述环路组织提出了一个模型,在该模型中,基底节处理不同区域的神经元表现出特定的解剖组织模式,高选择性地与下游进行通信。目前尚不清楚这些环路本身在多大程度上进一步与行为功能、执行协作以及学习过程中的可塑性相关。在许多情况下,同时选择或解除对两个或多个运动组件的抑制以执行动作非常重要,例如同时移动和重新定向。因此,同一环路必须适应协同运动的同时发生,要根据情境去区分并整合不同的运动模块。
下一节我们将介绍在运动调控、运动学习的过程中,基底神经节和脑干两个系统如何相互通信,并与其它运动相关网络(运动皮层和小脑输出神经元,以及多巴胺能神经元)集成,敬请期待!
聚精“汇”神系列科普活动
运动与我们的生活息息相关,尤其是最近挥洒汗水,但手忙脚乱的可爱刘畊宏男孩、女孩![]() ,一定意识到“有时候这手、脚就是不听使唤呐”!
,一定意识到“有时候这手、脚就是不听使唤呐”!![]() 为了让大家对参与运动的神经环路有更多的了解,AiBrain神经科学新媒体联合中国神经科学学会、上海神经科学学会、上海科协等机构,邀请南京大学生命科学学院副院长朱景宁教授,为我们讲述“运动与脑科学”,欢迎扫码观看!
为了让大家对参与运动的神经环路有更多的了解,AiBrain神经科学新媒体联合中国神经科学学会、上海神经科学学会、上海科协等机构,邀请南京大学生命科学学院副院长朱景宁教授,为我们讲述“运动与脑科学”,欢迎扫码观看!



声明:脑医汇旗下神外资讯、神介资讯、脑医咨询、AiBrain所发表内容之知识产权为脑医汇及主办方、原作者等相关权利人所有。未经许可,禁止进行转载、摘编、复制、裁切、录制等。经许可授权使用,亦须注明来源。欢迎转发、分享
