



如果你只是一个不经意的观察者,那你可能从外貌和行为上都区分不出一个动物群体中的单个成员。但仔细观察你会发现,动物根据它们与同伴的支配关系过着丰富多彩的社会生活。埃贝在1922年的论文里描述了鸡的优势等级——他称之为“啄序”,这一见解在上个世纪引发了社会等级研究的爆炸式增长。100年以后,社会等级已经成为生物学许多领域的关键组成部分。为了纪念“啄序”发现一百周年,我们AiBrain特别整理了社会等级领域一系列的重要综述与研究,以飨读者。本篇为啄序系列的第一篇综述。
动物之间存在着自发组织形成社会优势等级以规避攻击并提高整体生存的现象。
截至目前神经科学家们已经采用了几种模式生物(包括鱼类、爬行动物、啮齿类和灵长类动物) 来研究实验室环境中的优势等级,相关文献揭示了大脑如何表征社会等级从而指导动物在优势等级中做出适当的社会行为。那么,究竟有哪些大脑核团和功能脑区参与其中,它们分别扮演何等角色呢?
2022年1月10日,Salk研究所大牛Kay Tye等人在Philosophical Transactions of the Royal Society 杂志上发表了题为: Neural systems that facilitate the representation of social rank的综述,带我们走进动物优势等级行为背后的世界。

这篇综述从以下三个方面展开了讨论:
首先,作者讨论了大脑如何响应社会地位信号。
然后,作者讨论了社交接近和回避的学习机制,这些机制在作者认为可以推动产生适合等级的行为。
最后,作者讨论了大脑如何代表个体记忆(社会记忆),以及以上记忆如何支持在社会群体中维持独特的个体关系。
本文是主题为 “啄序百年:优势等级研究的现状和未来前景” 的一部分。
优势等级是在许多社会物种中发现的一种重要的社会组织形式,一些文章已提出攻击行为的神经机制是优势地位的一个重要特征,但对于社会等级在大脑中如何表现知之甚少。
有大量的行为证据表明,动物知道自己和其他群体成员在群体中的地位如何。比如,研究人员在从人类到鱼类的不同物种中观察到了“等级注意”,或者说低等级对更高等级的监控以避免攻击、从成功个体中观察学习,以及识别晋升机会。重要的是,在许多物种中,社会等级并非遗传,而是通过社会经验建立的,这表明学习机制对于社会等级的形成是必要的。动物评估附近同种动物社会地位的能力,是在适当的情境下表达主导或从属行为,进而维持稳定的社会地位的关键一步。在这篇综述中,作者探讨了与鱼类、爬行动物、啮齿动物和灵长类等社会物种的社会等级(尤其是类群中具有保守功能的机制)相关的神经回路(图1)。
作者首先回顾了参与状态信号的感知的下丘脑、中边缘和皮质回路。
然后作者表明负责探测信号的大脑区域与参与更一般社会学习过程的大脑区域互相重叠,并推测它们支持社会等级关系学习的机制,而这是一个相对未被研究的过程。
最后,作者探索了基于个体去识别社会等级的证据,以及可能支持群居物种之间更精细和认知更复杂的社会等级表征的机制。
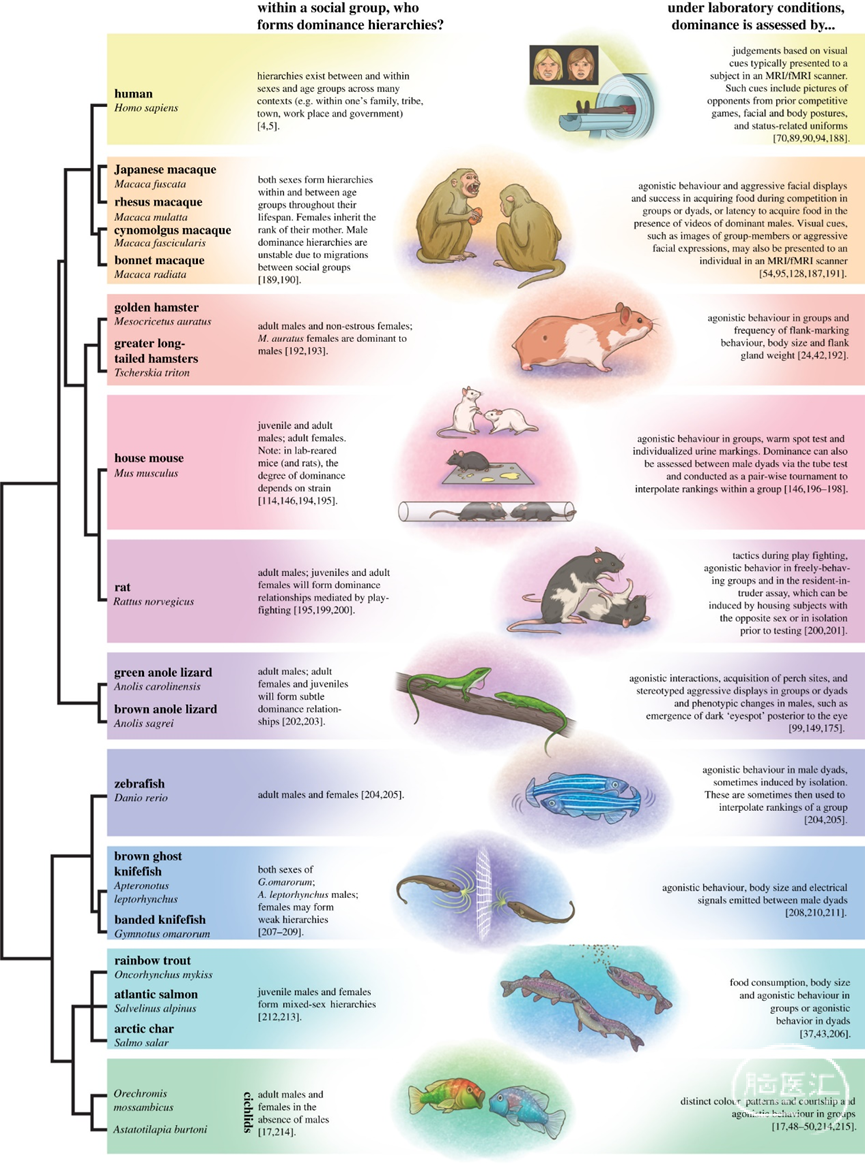
图1: 用于在实验室研究社会优势的常见物种。鱼类、蜥蜴、啮齿动物和灵长类动物在实验室环境中被用于研究优势和从属行为的神经基础。左边列出了记录等级的生命阶段和性别。右图显示了如何在实验室条件下评估优势。
优势或从属行为的表达并不总是需要体能竞争,而是可以基于个体竞争能力的特定线索,即地位信号。有人推测,地位信号进化用来在同物种之间传递关键信息,因此这些信号为优势个体提供了优先获得领土、资源和配偶的机会,同时也有助于避免从属个体的攻击。这些信号通常与信号传递者的成本有关,例如雄激素的产生或为增强其真实性而增加的被捕食的风险。重要的是,地位信号并不是任何已知的社会等级给定的社会刺激,而是在无论是否熟悉同种个体的情况下传达竞争能力的特定物种特征。地位信号在许多物种中都有广泛的应用,从信息素到复杂的行为表现,信号发起者也通常采用多种信号形式。因此,对社会地位的感知通常比检测单个信号更复杂,因为它涉及到多个特征的整合。尽管对大脑如何处理地位信号的理解有限,但通过研究涉及呈现化学和视觉信号的实验,研究者们发现了几个大脑区域和环路与处理地位信号有关(图2)。有趣的是,其中几个大脑区域也与一般的回避和奖励学习和记忆过程有关,因此作者还推测地位信号的表达到底是先天使然还是通过学习习得。
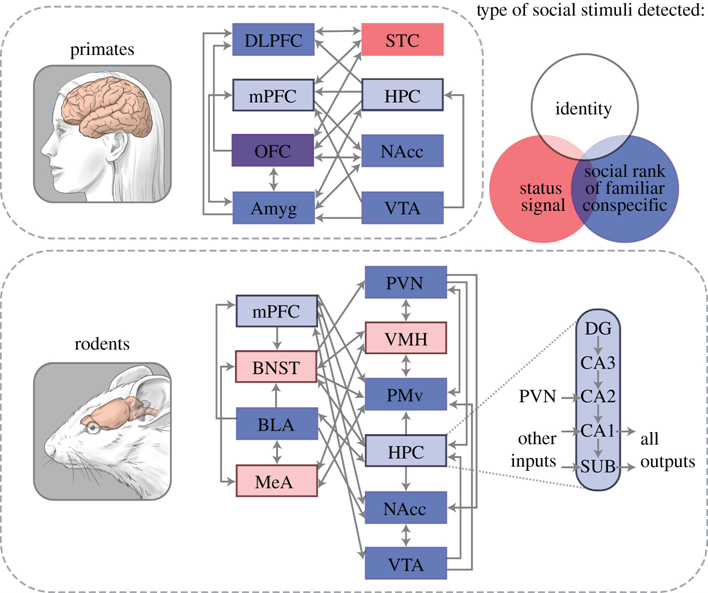
图2: 大脑系统参与表达优势等级。
(i)泛杏仁核和下丘脑回路处理啮齿类动物的化学信号
化学信号,例如信息素和尿蛋白,可能是社会物种用来传达包括性别、物种、地位和个体身份在内的识别信息最常见的交流方式。化学信号的使用在有地域性的物种中可能尤其重要,例如啮齿动物。在结合主嗅觉(挥发性气味)和犁鼻器/辅助嗅觉(非挥发性气味)系统的小型哺乳动物中,检测社交气味的神经回路被广泛研究。犁鼻器 (VNO) 神经元主要投射到与社会行为有关的区域,经典观点认为犁鼻系统专门用于检测与性别、生殖或优势地位信息有关的物种特异性化学信号,但有一些证据表明主嗅觉系统也会处理社会化学物质。在雄性小鼠(Mus musculus)中,VNO 中的特定受体是性别信息辨别,性行为和攻击性行为表达所必需的。在雌性和雄性小鼠中,VNO 的下游目标在嗅闻有等级优势的雄性尿液时比从低等级雄性的尿液中激活更明显,这可以通过 c-fos 免疫反应细胞的数量(神经元兴奋的代表)来证明。这种激活可能代表检测到更高水平的主要尿蛋白 (MUPs),在这其中有一些对于区域标记至关重要。此外,在啮齿动物中,特定的 MUP 代表着优势信号。
VNO的其中一个下游区域是内侧杏仁核(MeA),它接收来自犁鼻器和主嗅觉系统的输入,是脊椎动物气味处理的早期节点。MeA也是一个富含性类固醇激素受体的高度性二态核,与啮齿类动物、鱼类和蜥蜴的性行为和攻击行为有关。性别差异似乎也会影响MeA代表社会信息的方式。雄性和雌性小鼠MeA中的钙成像显示,神经元群和单个神经元的Ca2+动力学以性别特异性的方式区分同种线索,这表明生物相关性影响MeA中的社会气味表征。此外,神经肽催产素(OT)是一种已知的调节多种社会行为的物质,随着雄性动物性成熟,它促进了MeA在性识别中的作用,这表明MeA中的OT用于编码生物相关的气味。OT如何调节MeA中的社会地位信号处理尚不清楚,作者假设OT促进MeA中相关社会地位气味线索的编码。有趣的是,在雄性小鼠中,在受试者暴露于α(即等级中最具优势的个体)尿液后,MeA比次级同种小鼠有更多具有c-fos免疫反应性(cfos ir)的神经元,这并不取决于受试者自身的社会地位或他们对信息来源的熟悉程度,尽管这些因素可能会影响信息的生物学相关性。在对抗性遭遇后,优势和从属大长尾仓鼠(Tscheskia triton)的MeA活性也增加,而与攻击或防御行为的表达无关。这些发现可能表明,该区域的活动在总体上代表了感觉输入或唤醒,并且对于表达环境适当的主导或从属行为可能不是至关重要的。然而,有一些证据表明,MeA在化学感觉处理中的作用更加精细,因为MeA中的OT传递可能是编码个体同种记忆所必需的。
虽然MeA在根据小鼠尿液中的嗅觉信息来确定社会等级方面起着关键作用,但尚不清楚这一过程是否会被社交记忆进一步改变。有趣的是,一些MeA下丘脑投射靶点在小鼠接触尿液时被激活,并受社会地位和对尿液来源的熟悉程度的调节。例如,当最低等级的小鼠暴露于α尿液时,腹内侧下丘脑核(VMH)内的 c-fos ir差异比暴露于次级尿液大,无论尿液是否来自熟悉的小鼠。然而,只有当α小鼠暴露于不熟悉的α尿液时,VMH中的c-fos ir比熟悉或不熟悉的次级尿液中的c-fos ir更大。因此,接触尿液信息后VMH中c-fos ir受对线索的熟悉程度、提供信息的动物的社会地位以及处理信息的动物的影响。此外,在优势和从属动物中,当动物暴露于熟悉的α尿液时,与熟悉的从属尿液相比,腹侧乳头前核(PMv)表现出更大的c-fos ir。有趣的是,当动物暴露于不熟悉的α尿液和不熟悉的次级尿液时,PMv中的c-fos ir差异没有观察到。这表明,PMv没有检测等级信息的特征。相反,PMv似乎可以检测与社会等级记忆相关的特征。
综上证据表明,不同的大脑区域代表不同级别的社会等级。一些脑区比如MeA,似乎可以检测到地位信号,因为它们对优势和从属信息的反应表现出不同的c-fos ir,而不管是否熟悉。其他脑区,如VMH和PMv,则与包括动物自身的社会地位和对提供线索的个体的熟悉程度等有关。
(ii)其他物种的化学状态信号
在啮齿类动物之外,用于处理社会化学感觉信息的神经回路的特征不太明显,尽管社会化学物质似乎在视觉模糊的环境中对于其它类群特别重要,例如夜间活动的灵长类动物和生活在浑浊环境中的水生动物。与啮齿类动物一样,优势雄性慈鲷(Astatotilapia burtoni和Oreochromis spp.)使用尿液来表示优势地位,其尿液储存量比低等级要多。当池水被更新,分泌的化学物质被清除时,由于低等级在环境中对优势个体展开错误攻击,配对的鱼无法形成稳定的优势关系。有趣的是,当优势等级的雄性慈鲷暴露于另一个优势等级雄性的化学地位信号时,它们会增加排尿频率和循环的雄激素水平,而嗅觉加工区的基因表达模式与暴露于各种尿液信息有关。当优势等级雄性暴露于优势等级和低等级不同的尿液气味时,它们在嗅球(OB)和背端脑后部(Dp;哺乳动物嗅觉皮层的假定同系物)中也表现出不同的基因表达谱。此外,来自鱼类腹侧端脑腹侧核(Vv;与哺乳动物的侧隔(LS)和纹状体外苍白球同源)的神经记录表明,在这些区域,占优势的雄性更容易区分与性别和食物相关的气味,而低等级雄性的大脑活动只区分了占主导地位的雄性气味。因此,当暴露于社会信息时,鱼类的社会地位是影响它们如何处理这些信息的一个因素,在小鼠身上也观察到了这一点。
与鱼类和啮齿类动物不同,灵长类动物在评估社会信息时通常依赖视觉信息,而不是化学或嗅觉信息。然而,对夜间活动的新大陆猴子(如狐猴科、盲蝽科、盲蝽科和长尾猴科)的行为研究表明,生物气味可能参与与优势地位相关的活动,如区域标记和繁殖行为。目前对灵长类大脑如何处理社会嗅觉信息的研究有限,,然而有研究表明,副嗅球(AOB)的大小与社交和交配系统相关,因此社交网络更分散的物种具有更大的AOB容量,这表明在夜间活动的新大陆猴子中,类似于啮齿类动物的嗅觉通路被用于处理地位信号。灵长类是否能从化学信息中检测出社会地位仍然有待进一步研究,而关于灵长类动物大脑如何表示地位信号的大部分已知信息来自于使用视觉刺激的实验。
(i)灵长类动物视觉状态信号的杏仁核处理
对优势等级神经相关性的一些最早研究表明,杏仁核是大脑中适当表达优势和从属行为的关键区域。例如,在优势等级中,等级最高的猕猴(猕猴属)的双侧杏仁核损伤会对优势和从属行为以及社会等级的变化产生剧烈但可变的影响。这包括个体的攻击性完全降低,失去较高的地位,另一个个体的攻击性增加以及从典型的优势行为过渡到专制统治。人类病变研究,例如Klüver–Bucy综合征的研究,进一步支持杏仁核在情境下做出适当社会行为中的广泛作用。此外,在人类和非人灵长类动物中,个体的社交网络复杂性与杏仁核体积相关,这表明杏仁核在社会环境中起着导航作用。
杏仁核对于编码和传递包括社会刺激在内刺激所引起的情感效价至关重要。电生理学和神经成像研究表明,当猕猴和人类看到不熟悉的愤怒或威胁的个体的面部表情时,杏仁核的活动增加。在人类和猴子中,单个个体图像中愤怒的面部表情和直接的眼神接触与优势等级地位有关,因为这表明了对感知者的攻击。相反,猕猴露出的恐惧表情与顺从或缺乏攻击性有关。有趣的是,在人类中,杏仁核中诱发的血氧水平依赖性(BOLD)信号在面对恐惧面孔时反应更大,这表明不明来源的威胁会更强烈地激活杏仁核,而这支持了杏仁核的主要作用是获取环境中显著的社会信息这一观点。
(ii)杏仁核在学习社会等级刺激中的潜在作用
动物天生识别地位信号或后天学习所占程度尚不清楚。通常来说,检测信号和做出响应的过程是先天的,但这可能不适用于所有类型的信号。例如,尽管并没有多少警告性的面部表情,新生猴子杏仁核损伤依然会增加社交互动中的社交恐惧,而成年猴子的损伤则不会产生这种影响,反而会导致对陌生个体的谨慎行为减少。这些研究表明,要有效地解释特定的行为和信号,需要一段时间的社交和学习。此外,在没有状地位信号的情况下,杏仁核的活动与熟悉个体的社会等级相关。例如,人类的神经影像学研究表明,当受试者看到熟悉的主要竞争对手的中性面部表情时,杏仁核中的 BOLD 信号会增加,而猕猴基底外侧杏仁核 (BLA) 的放电率与中性熟悉面孔的社会等级相关。这表明杏仁核也可能代表与优势个体相关的社会等级学习或情感记忆。
在巴甫洛夫条件反射实验下,已经在啮齿类动物中彻底研究了杏仁核在恐惧和威胁学习关联中的作用。虽然条件反射实验传统上证明了电击与物体或环境之间的联系,但通过竞争产生的身体伤害很可能以类似的方式与地位信号相关。简而言之,恐惧条件反射过程的基础环路涉及BLA,BLA将有关外部环境和动物内部状态的感官信息与海马(HPC)和前额叶皮质(PFC)的上下文信息和记忆结合起来。这是学习引起恐惧的刺激(即非条件刺激:US)和中性刺激(即条件刺激:CS)之间关联的地方,然后这些信息被传递到中央杏仁核(CeA),中央杏仁核投射到中脑参与行为输出的区域。有趣的是,当啮齿类动物直接学习恐惧联想以及当它们通过观察同种动物(即替代学习)学习时,这个环路就会被激活,PFC会提供额外的自上而下的输入,以便在替代学习期间保持注意力。低等级动物可以通过观察群体成员之间的对抗性互动(使用替代学习)来确定社会等级,避免与直接学习相关的潜在伤害。虽然将同物种竞争能力与地位信号联系起来的联想学习机制可能是杏仁核地位信号表达的基础,但这一点尚未得到实证证实。
(iii)多巴胺能信号和腹侧纹状体对视觉状态信号的反应
在众多物种中,多巴胺能系统参与处理社会地位信号。在慈鲷中,雄性优势鱼的出现与腹侧端脑(Vc)中央区域的多巴胺(DA)神经元数量增加有关,腹侧端脑(Vc)是哺乳动物纹状体的同系物。在绿色无尾蜥蜴(Anolis carolinensis)中,特定脑区的DA信号在不同脑区升高,以响应地位信号的变化。用隐蔽优势信号眼点观察对手的雄性,其腹侧被盖区(VTA)、黑质(SNR)、伏隔核(NAcc)和下丘脑的DA增加,并成为优势。相比之下,用人工眼点观察对手的雄性蜥蜴中缝背核(DRN)和杏仁核的多巴胺(DA)增加,并成为低等级。
多巴胺能神经元强烈支配的病变区域似乎也会损害地位信号检测。腹侧纹状体单侧受损的蜥蜴无法检测到来自入侵的同物种的优势信号,这些信号出现在与受损半球相对应的视野部分,但当它们出现在完整半球时,没有发现损伤。有趣的是,人类(智人)腹侧纹状体的损伤也会削弱识别不熟悉的愤怒面部表情的能力。
除了信号检测之外,DA的释放还可以作为一个预测错误信号,提醒人们注意信号的经验值与其预期值之间的差异。这种现象建立在巴甫洛夫学习系统的基础上,因此DA信号可以将强度和效价分配给CS-US的关联,并促进强化学习行为模式,如社交接近和回避。与杏仁核中地位信号的表现类似,目前尚不清楚对地位信号做出反应的多巴胺能信号是先天的还是通过社会经验习得的。灵长类动物的研究表明,对优势等级刺激响应的DA信号增强是一种习得性反应。例如,人类的神经成像显示,腹内侧纹状体从多巴胺能输入接收有关奖赏的信息,当受试者看到熟悉的具有中性面部表情的优势个体的脸时,该区域显示出更大的BOLD信号。猕猴的电生理记录还揭示了内侧纹状体中的神经元群,这些神经元专门传递社会信息,在观察优势个体时,这些神经元的活动会增强。此外,雄性猕猴会牺牲果汁奖励,以便有机会看到熟悉的高等级猴子的脸。尽管这些研究没有直接记录多巴胺信号,但这些发现表明,DA信号的升高可能反映了观察优势群体成员的更大价值,因为这些提供了高等级动物的威胁信息。未来的研究使用最近开发的多巴胺成像传感器或许可以揭示这个假设。
(iv)灵长类动物视觉状态信号的皮层处理
考虑到PFC在社会认知和决策中的突出作用,如果多个前额叶区域对社会地位信号做出反应并不奇怪。例如,与非社会性刺激相比,眶额皮质(OFC)对同物种个体刺激图像的反应表现出更大的BOLD信号。此外,OFC的放电频率区分了恒河猴中常见的优势等级和低等级同种动物的图像。OFC内的活动也与社会形象价值相关(通过观看图像的时间来衡量)。然而,目前尚不清楚OFC是否代表习得的身份识别或某些优势相关的面部特征,因为本研究中的个体仅接触熟悉的面孔。对人类的研究表明,OFC对于识别面部表情和陌生受试者声音中的情绪非常重要。此外,神经影像学研究表明,背外侧前额叶皮质(DLPFC)在当受试者观察优势面部表情和熟悉的优势个体的中性面部时有更强的BOLD信号。
前额叶对地位的表达也可以反映注意过程和观察优势个体的显著性。这可以解释为什么当受试者观察优势面部表情和熟悉的优势个体的中性面部时,会观察到类似的BOLD信号反应。PFC在维持注意力方面的作用已被广泛研究。在对优势信号的反应中观察到的PFC活性升高可能表明地位信号有助于将一组动物的注意力指向优势焦点个体,这可能表现为一种注意力层次。有趣的是,认为自己地位低下的人类参与者对面部优势信号更敏感。PFC介导的注意力的这种潜在倾斜可能有助于获得更高等级的社会信息,即哪些群体成员具有高度的战斗性。
除了PFC,顶叶皮质区域也参与处理视觉状态信号。例如,当人类受试者呈现主要的面部特征和姿势时,以及当他们使用这些特征判断其他两个人的相对优势时,上颞叶皮质(STC)表现出更大的BOLD信号。也有证据表明,STC与颞下回或梭形回以及HPC一起工作,以收集面部反映的情绪和意图以及面部的个人身份信息
地位信号并不是动物感知社会地位的唯一方式。即使是依赖地位信号的物种,似乎也会优先考虑在竞争性互动中表现出优势地位的行为,以及对这些互动的记忆而非地位信号来指导它们的行为决策。例如,在蜥蜴身上,操纵优势信号暗眼点不会改变之前建立的主导-从属关系,但它确实决定了不熟悉的两个个体之间的主导-从属关系。此外,地位信号无法解释动物如何能够检测到存在于高度线性优势等级中的微妙等级梯度--其中个体对其两个最接近的等级组成员的行为也会存在差异。例如,来自同一层级的低级别小鼠的尿液气味特征没有明显的地位驱动差异。作者认为,动物还必须通过直接体验和与竞争对手的互动,观察其它个体之间的竞争(观察或替代学习),学习社会等级,以及如何与特定群体成员进行适当的行为,或者基于认知过程(如传递性推理)来最终确定社会地位。
值得注意的是,这些学习过程和地位信号并不相悖。此外,压力事件如优势信号的呈现,可以提高应激激素和神经调节剂的水平,从而增强学习和记忆。压力反应性和与地位信号的关系以及社会地位相关的神经内分泌特征已阐述详尽,本综述中不再讨论。在接下来的部分中,作者将讨论其推测的支持竞争能力和个体之间的联想学习的神经机制。
(a)杏仁核回路有助于学习社会从属关系和警惕性
作者认为杏仁核有助于通过竞争性社会互动学习社会等级关系。随着优势关系的建立,从属动物向更高级别的群体成员屈服,并避免因战斗而引起的疼痛和伤害,这种现象可能涉及恐惧和威胁学习系统。使用社会失败压力(SDS)范式对啮齿类动物进行的研究可能支持这一理论,但值得注意的是,在SDS期间,动物会持续进行对抗性互动,条件性社交失败的主要衡量标准是动物的社交互动倾向降低。SDS研究表明,杏仁核中类似的神经相关性是社交顺从和恐惧学习的基础。例如,脑源性神经营养因子(BDNF)介导的可塑性和BLA内的GABA能传递对于啮齿类动物条件性恐惧和条件性社交失败的获得和表达至关重要。内侧PFC(mPFC)向杏仁核的投射也显示出类似地调节恐惧条件反射和对SDS的反应。
BLA同系物在非哺乳动物中的作用也已被探索。在鳐鱼中,内侧背侧端脑(Dm)被认为与BLA同源。Dm与情绪学习和恐惧回避有关,通过c-fos免疫反应细胞数量反映的局部活动在SDS的后续几天增加,这表明它还介导了社交失败的学习和社交回避的发展。
虽然与恐惧和社交回避有关的杏仁核环路可能是一个群体中最低等级的动物如何学习其从属地位的基础,但它并不能解释等级中等的动物如何学会灵活地回避和服从某些群体成员,并对其它成员表现出攻击性。由对同种动物的恐惧学习或条件性社会失败引起的普遍性社会回避会阻碍中间动物在适当的时候表现出顺从和主导行为,并降低社交依恋行为。另一种解释中等等级灵活行为的可能性是,自然环境中的社交失败会导致类似焦虑的社会状态,比如社会警觉,而不是完全回避。社会警觉,或对同种动物的监测,可以加强观察学习和选择性地避免攻击同种动物。虽然高警觉被认为是对不确定性的一种不适应反应,并与焦虑症的高风险相关,但适当程度的警觉被认为是低级别动物采取的一种适应性应对机制,因为可以使其避免潜在的威胁,并抓住机会提高等级或获取资源。对哺乳动物的研究表明,作为扩展杏仁核的一个组成部分,终纹床核(BNST)在模糊的社交环境中传递刺激效价的,从而介导社交焦虑和社交警觉。在人类中,当参与者暴露于信号和威胁性面部表情之间不可预测的关联时,BNST的活动大于杏仁核的活动。
社交警觉似乎也受到自身社会地位的影响。在灵长类动物、啮齿类和鱼类中,社交警惕性是倾斜的,由此产生了等级注意,其中低等级动物的社会警惕性水平较高。在许多物种中,应激激素水平与社会地位之间存在负相关,但是在优势个体中也观察到高水平,揭示了应激激素与社会地位之间的U形关联。大多数研究都集中在低等级或被社交受挫的动物对对抗性社交互动的压力反应上。在小鼠SDS后,来自室旁核(PVN)的应激相关神经肽促肾上腺皮质激素释放因子(CRF)的释放上调。有趣的是,BNST中的CRF拮抗作用可以阻断仓鼠的条件性社交失败。这些发现支持了一个模型,在该模型中,应激激素介导了社交失败学习所需的BNST加工过程。
除了应激激素外,OT信号还根据个体的社会地位而变化,并调节社交警惕性。在暴露于压力下的雌性猕猴中,占主导地位与脑脊液中OT的升高有关。在雄性大鼠中, BNST和中央杏仁核(CeA)中的OT受体(OTR)在更具攻击性的品系中结合更高。在雄性小鼠中,OTR结合与其他扩展杏仁核(如NAcc和LS)中的优势社会地位相关。有趣的是,雄性小鼠BNST中的OTR敲除可防止由社会压力所引起的社交警惕性增加和社交方式的减少。在雄性猕猴中,OT的药理作用导致亲社会行为增加,并降低了受试者观察优势面孔的警惕性。总的来说,这些数据表明,与BNST和社会等级相关的应激激素和OT水平差异的机制可能会推动低等级动物的社交警惕性,进而形成等级注意。
虽然在非哺乳动物物种中,BNST与社会行为相关的功能尚未得到彻底研究,但最近的鱼类证据表明,腹侧端脑(Vs)的连合上亚区的活动被认为与MeA/BNST复合物同源,与非社会条件相比,在社交(即集体生活)中的信号学习过程中增加。因此,BNST在社会情境中的反应似乎在进化上是保守的。
(b)中边缘多巴胺系统介导社会支配和从属行为
多巴胺能系统是另一种可能参与社会等级学习的联想学习系统。当动物面对优势信号时,DA信号升高。此外,DRN中的多巴胺神经元以社会等级依赖的方式调节社会行为,此行为在更具优势的小鼠中激活DRN DA神经元的影响更大。虽然目前尚不清楚这是社会学习过程的结果,还是信号的先天反应,但有证据表明,DA信号可能在没有信号的情况下通过社会互动参与学习社会等级。
对啮齿类动物的研究表明,通过多巴胺能信号传导的奖赏学习介导攻击行为。在占优势的动物中,攻击性是一种自然的奖赏性US,可以强化攻击顺从动物的行为。在猕猴和小鼠中,优势雄性在纹状体中表现出更高的D2型DA受体结合,拮抗D2作用会减弱攻击性。在猕猴中,拮抗D2作用会导致优势等级的不稳定,尤其是对优势动物。一些研究还表明,D1和D2受体拮抗作用可以减弱大鼠和小鼠的攻击性。
虽然没有在优势等级环境中直接测试,但在啮齿类动物和灵长类动物中发现了一个高度保守的多巴胺能环路(图2),其中从VTA到NAcc(腹侧纹状体的一部分)的多巴胺能神经元促进社会奖赏的学习,并促进社会竞争力。值得注意的是,其他神经肽,5-羟色胺和OT,在小鼠的这个环路中对社会奖赏学习也很重要。鸟类、爬行动物、两栖动物和鱼类的VTA和NAc同系物之间也发现了类似的预测,它们激发了交配和求偶行为中的社交接近。
目前尚不清楚DA受体表达的差异是否预先决定了动物的社会地位,或者它们是否是后天社会地位差异的结果。例如,最初动机更强的动物通过获胜的奖励体验使这种行为得到更大强化。DA信号的增强与赢家效应有关,赢家效应是指获胜的战斗会使未来与新个体的较量持续下去,从而进一步提高社会地位。此外,当观看对手的图像时,具有高度竞争动机的人类表现出更高的腹侧纹状体激活。
有趣的是,DA信号也参与调解低等级动物行为。SDS模型类似于等级体系中下级动物经历的反复失败场景。通过调节可塑性,VTA–NAc环路中的DA传输与小鼠和仓鼠的社交受挫学习有关。此外,在社会性一夫一妻制啮齿动物中,SDS诱导MeA中D1受体的上调,而MeA是处理社会信息的重要区域。因此,SDS后DA信号的变化表明,DA传递和DA受体表达的个体差异可能是获得特定社会等级的结果,而不是预先存在的社会等级相关性。这些研究还表明DA信号的奖赏效应与厌恶效应存在大脑区域特异性。
总之,物种间的多巴胺环路促进了社会奖赏学习,并调节了动物与特定个体接近或回避的决定。这种学习机制可以用于动物发展社会等级,并在该等级中学习它们的社会等级。
生活在社会优势等级中的动物必须动态地表现行为,以便它们对某些个体表现出主导行为,对另一些个体表现出从属行为。因此,个体群体成员的社会记忆需要与促进接近和回避社会行为的联想学习过程相结合。在高度线性的优势等级体系中,每个个体都有一个独特的社会等级,动物的行为存在细微的等级差异。例如,占优势的雄性小鼠倾向于对排名靠前的竞争对手表现出更多攻击性,而雄性黑猩猩倾向于只为排名靠前的雄性梳理毛发。这种社会选择性行为表明,动物对其同种动物有很强的个体社会记忆。动物是否总是依赖于个体的社会记忆或使用与地位相关的概括特征(即地位信号),可能取决于社会环境的动态和选择压力。例如,生活在高度动态的社会环境中的雄性慈鲷会形成对优势对手的短期记忆,只会对在过去7天内表现出超强战斗力的具有优势形态的雄性表现出顺从行为。类似地,蜥蜴通过单一的社会互动形成社会记忆,但随着后续互动间隔的增加,这种社会身份信息的显著性似乎会下降,并最终被遗忘。此外,群居而非单居的小鼠即使在短暂的互动后也能表现出个体的长期记忆,这进一步支持了社会记忆依赖于社会背景的观点。接下来作者将回顾个体社会认知的神经基础,这可能有助于在优势等级中学习个体的社会等级。
(a)海马和PHC回路在社会记忆编码和回忆中的作用
广泛物种的研究认为HPC对情景记忆和语义记忆都至关重要。最近的研究探索了HPC在社会记忆中的具体作用,包括表征个体和社会地位关系。患有海马病变的人类患者无法识别熟悉的面孔。更具体地说,对啮齿类动物的研究表明,腹侧HPC(vHPC;灵长类动物的前HPC;aHPC)通过与下丘脑和杏仁核的连接调节动机行为,并在效价相关学习中发挥作用。与下丘脑和杏仁核的相互联系,以及vHPC具有记忆整合效价信息的能力,使该神经系统在社会记忆中发挥作用。在健康人类受试者中,当受试者面对同一个体以各种姿势和情境呈现时,aHPC中的神经元被激活,这表明不同的神经元群代表了个体的社会记忆。在小鼠中抑制vHPC会损害个体的记忆编码和回忆能力,在第一次社交接触中活跃的vHPC神经元的光遗传学再激活可以促进这些过程。
此外,最近对小鼠进行的研究发现,海马内回路与社会记忆编码和回忆有关。HPC由CA1、CA2、CA3和齿状回亚区组成(图2),并延伸至整个内侧颞叶。背侧CA2(dCA2;灵长类动物的后部)是啮齿类动物记忆同窝动物所必需的,从dCA2到腹侧CA1(灵长类动物的前部)的投射可以编码、存储和回忆社会记忆。在鱼类、两栖动物和爬行动物中发现了同源的海马结构和亚区,它们编码的空间记忆与哺乳动物相似,但对它们在社会记忆中的作用知之甚少。然而,在蜥蜴身上,当地位信号被操纵到与对对手地位状态的记忆不一致时,CA3的功能就会发生变化。此外,在鱼类中,与哺乳动物HPC同源的背侧端脑(Dl)外侧部分的亚区在社交环境下与非社交环境下表现出不同的c-fos免疫反应性。然而,一项研究表明,HPC是否在社会等级背景下的社会等级学习中发挥了作用尚未得到直接测试。BDNF可以增强学习所需的神经可塑性,在赢得竞争性互动后,优势小鼠HPC中的BDNF上调,这可能随后增强其优势社会地位。
在哺乳动物中,额叶皮质及其与HPC的连接也促进了特定群体成员记忆的编码和存储。对啮齿类动物的研究表明,vHPC对mPFC的投射对于回忆社会记忆是必要的。人类的神经影像学研究表明,mPFC和HPC BOLD活动都与游戏中学习社会等级相关。在人类中,头侧mPFC的经颅电刺激促进了社会等级学习。此外,在小鼠中,当与同种动物竞争时,mPFC群体放电频率可预测相对社会等级和绝对社会等级。
总之,有强有力的证据表明,HPC在编码个体社会记忆中的作用在社会物种中是保守的。然而,海马内回路的复杂性以及它们如何促进记忆形成主要是在啮齿类动物种得以研究。在哺乳动物中,社会记忆环路延伸到PFC,这是一个对长期记忆至关重要的区域。尽管人类的相关研究指出HPC–mPFC通路在社会等级学习中的作用,但这尚未在动物模型研究中得到证实。目前仍需要进一步研究非哺乳动物大脑中社会记忆的存储方式来确定个体识别是否是稳定社会等级的保守要求。
(b)神经肽支持社会记忆编码
众所周知,神经肽OT和精氨酸加压素(AVP)可调节不同物种的社会认知和行为,在支持社会记忆方面具有特殊作用。大多数AVP和OT合成神经元起源于下丘脑的视交叉上核(SCN)和PVN,广泛投射到整个大脑,而HPC是主要的投射目标之一。在大鼠中,海马AVP受体是编码个体同物种记忆所必需的,刺激AVP传入dCA2有助于社会记忆编码。在小鼠中,dCA2/CA3中的OTR对于同种动物的短期记忆和长期记忆都是必需的。
OT和AVP的传播还通过作用于杏仁核和扩展杏仁核的成分影响社会记忆。对小鼠和大鼠的研究表明,以处理社会气味而闻名的MeA还可编码个体的同物种记忆。在MeA中阻断OTR会损害啮齿类动物的社会认知,在初次接触时激活这些受体会增强它们编码社会记忆的能力。同样在啮齿类动物中,MeA和BNST向LS发送AVP投射,在LS中,AVP受体1(V1aR)的阻断或下调扰乱了社会认知。从MeA到vHPC的AVP投射也已在大鼠大脑中得到证实,但尚不清楚这些投射是否也与社会记忆有关。
尽管来自啮齿类动物和灵长类动物的证据强烈支持神经肽在社会记忆中的功能,但鱼类和蜥蜴中同源神经肽的功能仍有待确定。精氨酸血管催产素(AVT)是在鱼类和爬行动物中发现的AVP同系物,在非社交空间和信号学习中发挥作用,但其在社交环境和社交刺激学习中的作用尚未确定。值得注意的是,AVT与求爱行为、攻击性和其他等级相关行为(如通过尿液发出状态信号)的增加有关。AVP与哺乳动物的类似社会行为有关,表明其功能在很大程度上是保守的,支持研究其在跨物种社会记忆中的作用。类似地,在鱼类和爬行动物中分别发现的OT同系物鱼神经叶激素(isotocin)和 中催产素(mesotocin)在社交方式、性行为、伴侣关系的形成和养育行为方面似乎与OT类似。需要进一步研究AVP和OT同系物是否调节同种动物的记忆,以了解这些神经肽是否在跨社会物种的社会记忆编码中起到保守作用。
本篇综述详细讲解了社会等级识别涉及跨多个认知水平,并具有高度保守性的神经环路的协调活动,包括从看似天生的对社会地位信号的感知到对特定个体社会等级的更精细的学习,并绘制了人类和啮齿类动物参与优势等级神经网络图谱,探讨两栖类鱼类类似的神经机制。为之后更详尽的优势等级研究做出了重要总结和参考。
基于目前研究的有限性,该综述也进一步提出问题和思考方向:
虽然地位信号似乎有助于绕过经验学习以及避免可能导致伤害的社交互动,但地位信号反应是先天的还是后天习得的仍需要更彻底地研究。此外,个体的社会地位似乎会影响与获取社会信息相关的行为,如注意姿势和视觉注视方向,但社会信息在等级动物大脑中的差异表现尚不清楚。另外,尽管本综述中讨论的物种形成了优势等级,但参与社会等级表征的神经系统的证据通常并不直接来自在群体中自由生活和行为的动物。基于实验室的神经生物学和行为学研究中,简单的二元社会互动分析的比例过高,这些分析不具有群体中社会等级的代表性。传统上只测量男性表达的行为,因此本综述中介绍的大部分知识都来自几乎完全在雄性动物身上进行的实验,雌性动物是否具有相似的神经机制和行为学表现仍有待进一步探究。
在招岗位
2022 RECRUIT /
招募编译团队及审稿团队
岗位职责:撰写、审阅神经科学各领域的文献导读、领域速报。
岗位要求:神经科学领域四年级以上的博士生、博士后以及青年科研工作者。
AiBrain作者团队由海内外知名高校博士生、博士后,及已成立实验室的PI们组成;加入AiBrain,可以体验专业的投审稿系统、高效温馨的团队合作、丰富的文章栏目,以及有竞争力的丰厚报酬;能力突出者可以作为领域/栏目负责人,并配有岗位津贴。
表现突出者将以“AiBrain特邀记者”的身份,参加今年9月份在苏州举办的神经科学年会,对院士特邀报告等重要大会报告进行解读和撰稿。
欢迎有意者投递简历及个人作品!

微信号:AiBrainzhushou
邮箱:ai_brain@163.com
