




以往的研究表明,面对竞争性视觉刺激时,动物通常会从多个竞争刺激中针对一个对象做出行为反应,或者综合所有刺激做出反应,也就是“赢家通吃策略”和“平均策略”。其中,赢家通吃策略(Winner-take-all,WTA)指的是动物对单一目标做出反应而不考虑其他目标,被认为在自下而上、刺激驱动的注意过程中至关重要;与之相对的平均策略(Averaging),则会在整合了多个相互竞争的感官刺激后做出反应。但在2021年的《Neuron》杂志上,德国马普所的Herwig Baier实验室通过幼年斑马鱼这一模式生物,解析了视觉竞争背后的神经机制。Baier实验室之所以选择将幼年斑马鱼作为研究对象,是因为幼年斑马鱼有小而透明的大脑,可以进行活体全脑成像,并且还有丰富的行为,优势显著。
为了更好地定量分析,作者在实验过程中使用膨胀的阴影刺激(Looming)来模拟捕食者的靠近。他们发现,斑马鱼在面对刺激时,会混合地使用WTA和Averaging策略,前者约占80%,后者约占20%(图一)。
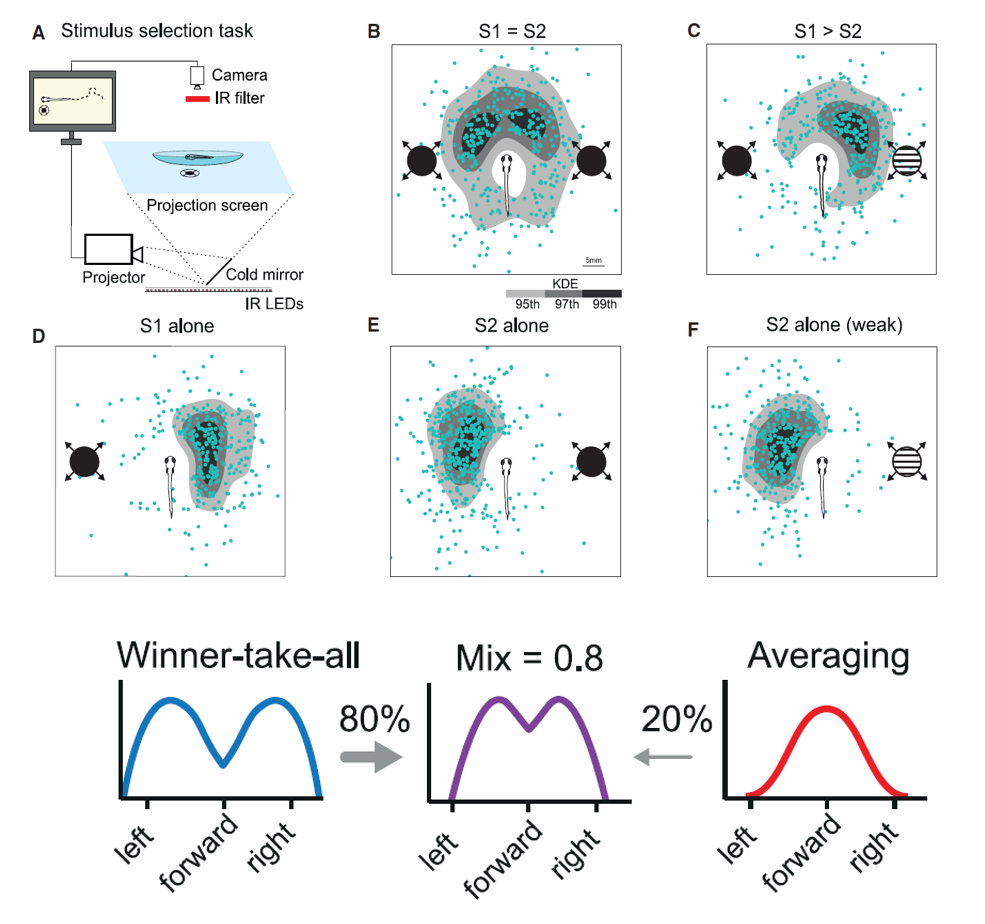
图一
在行为学上发现了如此明显的差异后,作者又利用全脑钙成像技术研究刺激选择的神经机制。作者推测,执行WTA策略,至少需要两种类型的神经元,一种是“和其中一个刺激强度反应一致”的神经元,另一种是“活动被另一个更强的刺激抑制”的神经元。
为了区分这两类神经元的位置和功能,作者设计了一个单眼刺激的行为范式,即保持刺激1(stimulus 1, S1)的膨胀速度不变,而让S2(stimulus 2, S2)的膨胀速度由弱变强。全脑钙成像的结果显示,在视网膜神经节细胞(retinal gangalia cell, RGC)的轴突和顶盖(tectal neuron)神经元上都记录到了预期的两种信号,第一类信号(绿色)随着S2的增强而增强,第二类信号(紫色)随着S2的增强而受到抑制。经过统计,作者还发现在顶盖神经元中信号发生改变的速率更快。(图二)
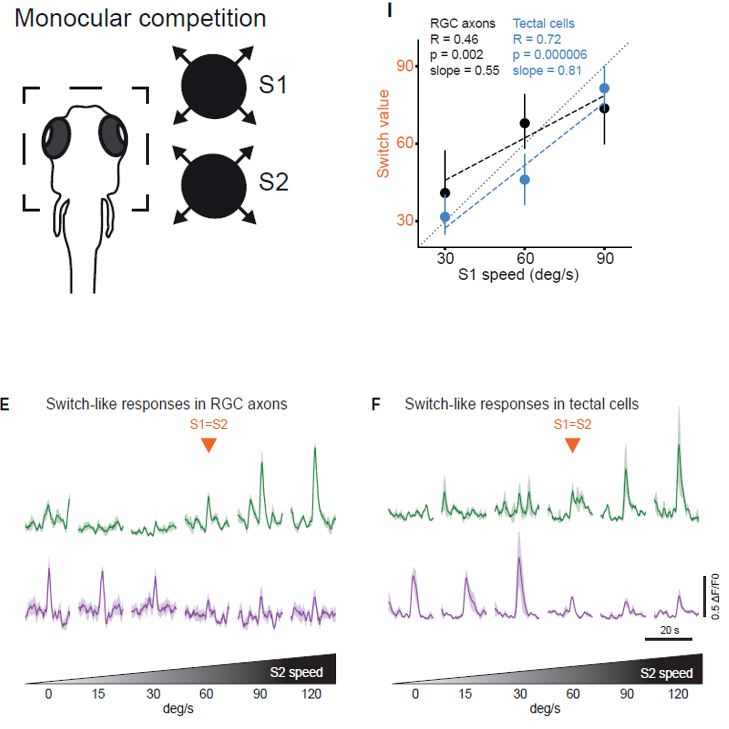
图二
我们知道,RGC是顶盖的上游。作者想进一步确认顶盖神经元对RGC是否有逆向调控,因此对顶盖神经元进行了损毁,结果发现RGC轴突的信号并没有受到影响;这说明RGC轴突的反应不需要顶盖神经元进行逆向调节。也就是说,这种信号先在RGC轴突出现,并且在顶盖神经元进一步增强。(图三)
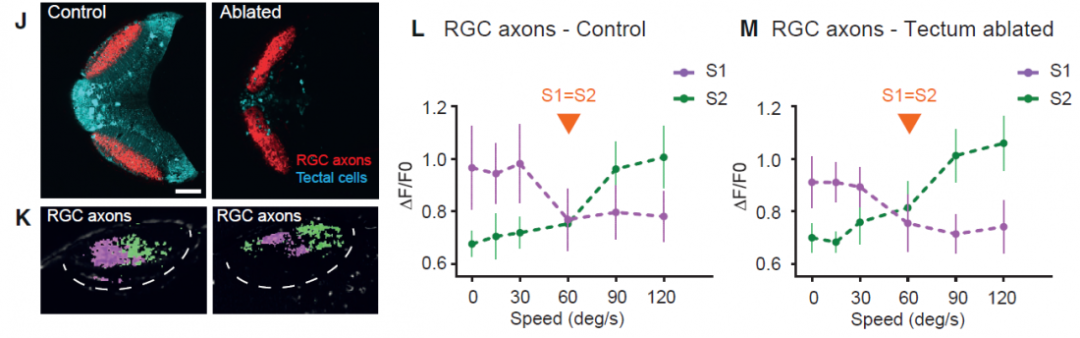
图三
既然视网膜-顶盖的WTA是用于刺激选择的通用机制,那问题来了:这一WTA策略是如何影响下游神经环路的呢?

事实上,科学家们早就发现,鸟类的NI核团参与视网膜对双眼竞争(binocular competing)的反应。于是,作者很自然地关注了NI核团的动力学性质。不出所料,NI神经元的变化与顶盖神经元十分相似。(图四)
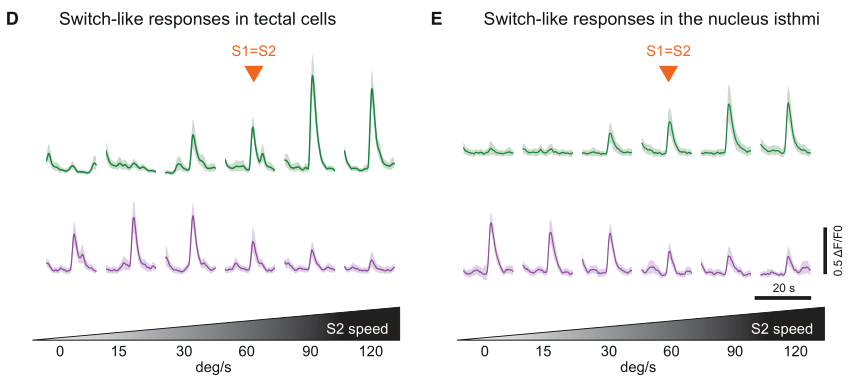
图四
为了进一步明确NI神经元的组成,作者使用了一些对神经递质特异的Gal4转基因鱼系,并了解到NI核团是由抑制性的GABA能神经元和兴奋性的谷氨酸能神经元组成。(图五)
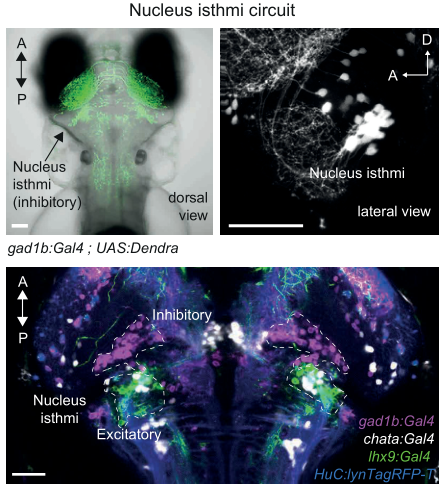
图五
为了探究某一核团对行为学的作用,生物学家通常会进行两方面的检验:功能获得性实验(gain-of-function)和功能缺失性实验(loss-of-function)。在神经科学领域中,功能获得性实验一般包括光遗传激活或化学遗传激活实验,而功能缺失性实验一般是指光遗传抑制、化学遗传抑制或损毁实验。
为了验证NI神经元对行为的影响,作者首先进行了功能获得性实验。他们期望改变NI神经元的活性,然后观察游泳行为的变化,于是,作者就在NI神经元中表达了光敏感抑制性蛋白GtACR2和光敏感激活蛋白ChR2,随后在looming刺激出现时抑制NI神经元的活性。结果表明,幼鱼的C-start逃跑行为的数目大大降低,而光遗传激活NI神经元则会直接减少游泳行为。这充分地说明了NI神经元会影响幼鱼的行为。(图六)
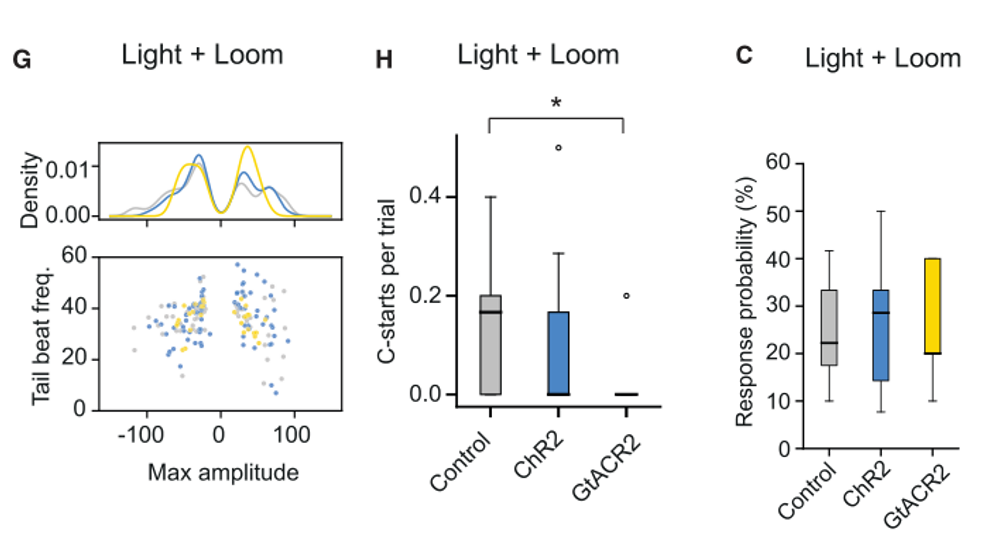
图六
既然存在行为学上的效应,那NI是否存在对顶盖神经元的投射?为了探究NI与顶盖之间的神经环路,作者又对NI中的兴奋性神经元进行了光遗传激活,并观察顶盖神经元的活性变化。结果显示,顶盖神经元的反应与受视觉刺激引发的反应很类似:既存在单侧顶盖神经元的反应(与WTA策略类似),也存在双侧顶盖的反应(与averaging策略类似)。不仅如此,在呈现looming刺激时,作者光遗传激活了NI中的8-10个细胞,发现NI和顶盖神经元的活性获得了不同程度的调节。这些都说明NI会调节顶盖神经元对looming刺激的反应。(图七)
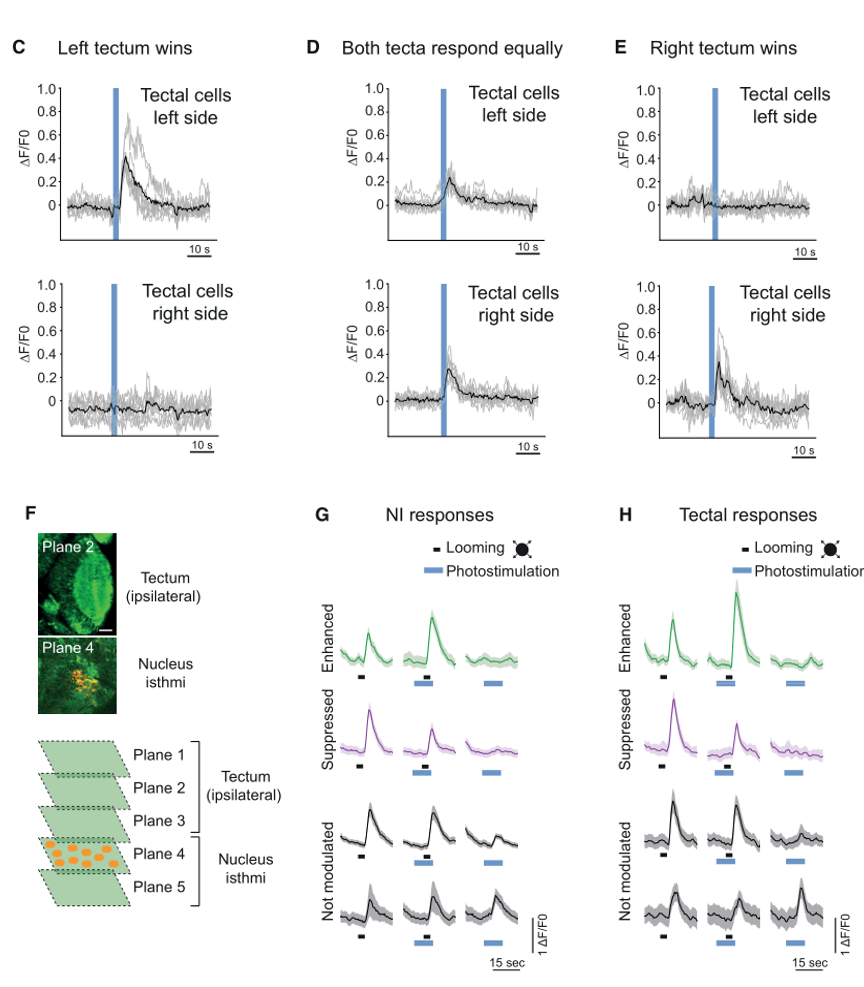
图七
为了进一步验证NI与顶盖之间神经环路的必要性,作者接着开展了功能缺失性实验。他们发现,双光损毁特定的NI神经元会破坏鱼对单边looming刺激的反应,损毁任意一侧的GABA能神经元都会降低顶盖对looming的反应,而损毁任意一侧的Lhx9+神经元都会增强顶盖对looming的反应;这与之前的结论一致说明了NI与顶盖之间存在神经环路的连接。(图八)
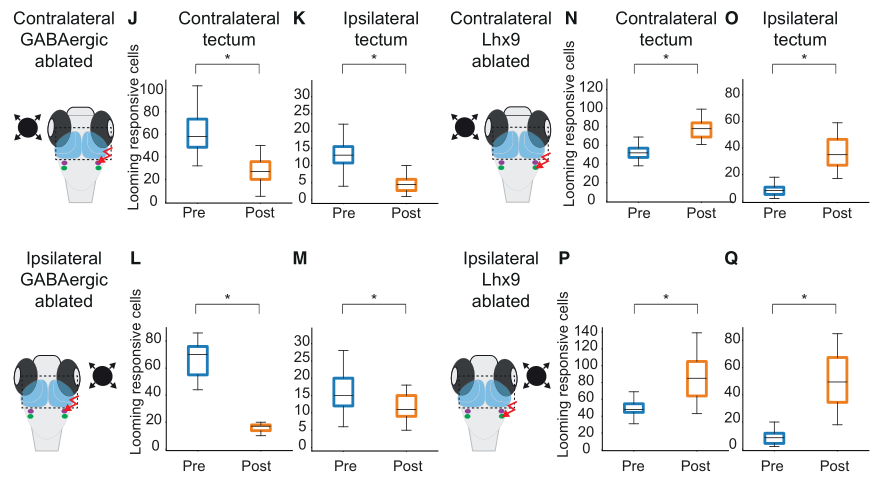
图八

本文在结构学研究的基础之上,采用了多种功能学操纵的手段,探究了幼年斑马鱼视觉竞争的神经机制,证明在单眼竞争时,神经系统通过交互抑制的方式,采取WTA的策略来激活对侧顶盖神经元;而在双眼竞争时,由于双眼接受到的刺激不同,引起的NI和顶盖之间的神经环路也存在差异。

1

声明:脑医汇旗下神外资讯、神介资讯、脑医咨询、AiBrain所发表内容之知识产权为脑医汇及主办方、原作者等相关权利人所有。未经许可,禁止进行转载、摘编、复制、裁切、录制等。经许可授权使用,亦须注明来源。欢迎转发、分享。
